Строение и функции ядра
Ядро (лат. nucleus, греч. karion-ядро) – это обязательный компонент эукариотических клеток. Оно хорошо различимо в неделящихся клетках и выполняет ряд важнейших функций:
1) хранение и передача наследственной информации в клетке;
2) создание аппарата белкового синтеза – синтез всех видов РНК и образование рибосом.
Выпадение или нарушение любой из этих функций приводит клетку к гибели.
Клетка содержит, как правило, одно ядро, но имеются двуядерные и многоядерные клетки.
Интерфазные ядра состоят из: ядерной оболочки, ядерного сока (кариоплазма, кариолимфа или нуклеоплазма), ядерного белкового остова, хроматина и ядрышек.
Ядерная оболочка (кариолемма) состоит из двух мембран, между которыми имеется перинуклеарное пространство шириной 10-40нм, заполненное электронно – микроскопически рыхлой субстанцией. Наружная мембрана ядерной оболочки со стороны цитоплазмы в ряде участков переходит в мембраны эндоплазматической сети, и на ее поверхности располагаются полирибосомы. Внутренняя мембрана ядерной оболочки участвует в обеспечении внутреннего порядка в ядре – в фиксации хромосом в трехмерном пространстве. Эта связь опосредуется с помощью слоя фибриллярных белков, сходных с промежуточными филаментами цитоплазмы.
В ядерной оболочке имеются поры диаметром около 90 нм. В этих участках по краям отверстия мембраны ядерной оболочки сливаются. Сами отверстия заполняются сложноорганизованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и заполняющих их структур называется поровым комплексом.
По краю порового отверстия располагаются в три ряда гранулы (по 8 гранул в каждом ряду). При этом один ряд лежит со стороны цитоплазмы, другой – со стороны внутреннего содержимого ядра, а третий – между ними. От гранул этих слоев радиально отходят фибриллярные отростки, образуя в поре как бы перепонку – диафрагму. Фибриллярные отростки направляются к центрально расположенной грануле.
Поровые комплексы участвуют в рецепции транспортируемых через поры макромолекул (белков и нуклеопротеидов), а также в активном переносе через ядерную оболочку этих веществ с использованием АТФ.
Число ядерных пор зависит от метаболической активности клеток. Чем интенсивнее протекают в клетке процессы синтеза, тем больше пор. В среднем на одно ядро приходится несколько тысяч поровых комплексов.
Основные функции ядерной оболочки следующие:
- барьерная (отделение содержимого ядра от цитоплазмы и ограничение свободного доступа в ядро крупных биополимеров);
- регуляция транспорта макромолекул между ядром и цитоплазмой;
- участие в создании внутриядерного порядка (фиксация хромосомного аппарата).
Кариоплазма (ядерный сок, или нуклеоплазма, или кариолимфа) – это содержимое ядра, которое имеет вид гелеобразного матрикса. Она содержит различные химические вещества: белки (в том числе и ферменты), аминокислоты и нуклеотиды в виде истинного или коллоидного раствора.
Ядерный или белковый остов (матрикс). В интерфазных ядрах негистоновые белки образуют сеть – «белковый матрикс». Он состоит из периферического фибриллярного слоя, выстилающего ядерную оболочку (ламина), и внутренней сети, к которой прикрепляются фибриллы хроматина. Матрикс участвует в поддержании формы ядра, организации пространственного положения хромосом. Кроме того, в нем содержатся ферменты, необходимые для синтеза РНК и ДНК, а также белки, участвующие в компактизации ДНК в интерфазных и митотических хромосомах.
Хроматин – комплекс ДНК и белков (гистоновых и негистоновых). Хроматин является интерфазной формой существования хромосом.
В этот период разные участки хромосом имеют неодинаковую степень компактизации. Наибольшей степенью компактизации обладают генетически инертные участки хромосом. Они хорошо окрашиваются ядерными красителями и называются гетерохроматином. Различают конститутивный и факультативный гетерохроматин.
Конститутивный гетерохроматин образован нетранскрибируемой ДНК. Полагают, что он участвует в поддержании структуры ядра, прикреплении хромосом к ядерной оболочке, узнавании при мейозе гомологичных хромосом, разделении соседних структурных генов и в процессах регуляции их активности.
Факультативный гетерохроматин, в отличие от конститутивного, может становиться транскрибируемым на определенных стадиях клеточной дифференцировки или онтогенеза. Примером факультативного гетерохроматина может служить тельце Барра, образующееся у организмов гомогаметного пола за счет инактивации одной из Х-хромосом.
Декомпактизированные участки хромосом, которые плохо окрашиваются ядерными красителями, называются эухроматином. Это функционально активный, транскрибируемый хроматин.
Ядрышки – уплотненные тельца, обычно округлой формы, диметром менее 1 мкм. Присутствуют они только в интерфазных ядрах. Количество их колеблется в диплоидных клетках от 1 до 7, но в некоторых видах клеток, например, микронуклеусах инфузории, ядрышки отсутствуют.
Ядрышки содержат более 80% белка и около 15% р-РНК, а также ядрышковый хроматин. Ядрышки образуются в области вторичных перетяжек хромосом, представленных ядрышковыми организаторами. При транскрипции этих участков хромосом образуется рибосомальная РНК, которая, связываясь с белками, участвует в формировании малых и больших субъединиц рибосом.
8 вопрос. ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
Цитоплазматическая мембрана или плазмалемма (лат. membrana – кожица, плёнка) – тончайшая пленка (7– 10нм), отграничивающая внутреннее содержимое клетки от окружающей среды, видна только в электронный микроскоп.
По химической организации плазмалемма представляет липопротеидный комплекс – молекулы липидов и белков.
Её основу составляет липидный бислой, состоящий из фосфолипидов, кроме этого, в мембранах присутствуют гликолипиды и холестерол. Все они обладают свойством амфипатричности, т.е. у них есть гидрофильные («любящие воду») и гидрофобные («боящиеся воды») концы. Гидрофильные полярные «головки» липидных молекул (фосфатная группа) обращены кнаружи мембраны, а гидрофобные неполярные «хвосты» (остатки жирных кислот) – друг к другу, что создает биполярный липидный слой. Молекулы липидов подвижны и могут перемещаться в своем монослое или редко – из одного монослоя в другой. Монослои липидов обладают ассиметричностью, т. е. отличаются по составу липидов, что придает специфичность мембранам даже в пределах одной клетки. Бислой липидов может находиться в состоянии жидкого или твердого кристалла.
Вторым обязательным компонентом плазмалеммы являются белки. Многие мембранные белки способны перемещаться в плоскости мембраны или вращаться вокруг своей оси, но не могут переходить с одной стороны бислоя липидов на другой.
Липиды обеспечивают основные структурные особенности мембраны, а белки – её функции. Функции мембранных белков различны: поддержание структуры мембран, получение и преобразование сигналов из окружающей среды, транспорт некоторых веществ, катализ реакций, происходящих на мембранах.
Различают несколько моделей строения цитоплазматической мембраны.
①. БУТЕРБРОДНАЯ МОДЕЛЬ (белки – липиды – белки)
В 1935г. английские ученые Даниэли и Даусон высказали идею о послойном расположении в мембранемолекул белков (темные слои в электронном микроскопе), которые залегают снаружи, и молекул липидов (светлый слой) – внутри. Длительное время существовало представление о едином трехслойном строении всех биологических мембран.
При детальном изучении мембраны с помощью электронного микроскопа оказалось, что светлый слой на самом деле представлен двумя слоями фосфолипидов – это билипидный слой, причем водорастворимые его участки – гидрофильные головки направлены к белковому слою, а нерастворимые (остатки жирных кислот) – гидрофобные хвосты обращены друг к другу.
 |
Однако уже с середины 60-х годов начали накапливаться факты против унитарной «бутербродной» модели. В частности, по одним данным, не все мембраны имели четкую трехслойную структуру при электронно-микроскопическом исследовании; по другим – значительная часть мембранных белков имела глобулярную структуру, а не ламеллярную, как в постулируемой модели. Наконец, среди многочисленных моделей мембран, предложенных в середине 60-х годов, начали выделяться те, в которых доказывалось наличие гидрофобно-гидрофильных взаимодействий не только между липидными молекулами, но и между липидами и белками.

|

 ②. ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ
②. ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ
В 1972г. Сингер и Николсон описали модель мембраны, которая получила широкое признание. Согласно этой модели молекулы белков не образуют сплошного слоя, а погружены в биполярный липидный слой на разную глубину в виде мозаики. Глобулы белковых молекул, подобно айсбергам, погружены в «океан»
липидов: одни находятся на поверхности билипидного слоя – периферические белки, другие погружаются в него наполовину – полуинтегральные белки, третьи – интегральные белки – пронизывают его насквозь, формируя гидрофильные поры. Периферические белки, находясь на поверхности билипидного слоя, связаны с головками липидных молекул электростатическими взаимодействиями. Но они никогда не образуют сплошного слоя и, по сути дела, не являются белками собственно мембраны, а, скорее, связывают ее с надмембранной или субмембранной системой поверхностного аппарата клетки.
Основную роль в организации собственно мембраны играют интегральные и полуинтегральные (трансмембранные) белки, имеющие глобулярную структуру и связанные с липидной фазой гидрофильно-гидрофобными взаимодействиями. Молекулы белков, как и липиды, обладают амфипатричностью и своими гидрофобными участками взаимодействуют с гидрофобными хвостами билипидного слоя, а гидрофильные участки обращены к водной среде и образуют с водой водородные связи.
③. БЕЛКОВО-КРИСТАЛЛИЧЕСКАЯ МОДЕЛЬ (модель липопротеинового коврика)
Мембраны образованы переплетением липидных и белковых молекул, объединяющихся между собой на основе гидрофильно-
гидрофобных взаимодействий.
 | |||
 | |||

Белковые молекулы, как штифты, пронизывают слой липидов и выполняют в составе мембраны функцию каркаса. После обработки мембраны жирорастворимыми веществами белковый каркас сохраняется, что доказывает взаимосвязь между молекулами белков в мембране. По-видимому, эта модель реализуется лишь в отдельных специальных участках некоторых мембран, где требуется жесткая структура и тесные стабильные взаимоотношения между липидами и белками (например, в области расположения фермента Na-К –АТФ-азы).
Самой универсальной моделью, отвечающей термодинамическим принципам (принципам гидрофильно-гидрофобных взаимодействий), морфо-биохимическим и экспериментально-цитологическим данным, является жидкостно-мозаичная модель. Однако все три модели мембран не исключают друг друга и могут встречаться в разных участках одной и той же мембраны в зависимости от функциональных особенностей данного участка.
СВОЙСТВА МЕМБРАНЫ
1. Способность к самосборке. После разрушающих воздействий мембрана способна восстановить свою структуру, т.к. молекулы липидов на основе своих физико-химических свойств собираются в биполярный слой, в который затем встраиваются молекулы белков.
2. Текучесть. Мембрана не является жесткой структурой, большая часть входящих в её состав белков и липидов может перемещаться в плоскости мембраны, они постоянно флюктуируют за счет вращательных и колебательных движений. Это определяет большую скорость протекания химических реакций на мембране.
3. Полупроницаемость. Мембраны живых клеток пропускают, помимо воды, лишь определённые молекулы и ионы растворённых веществ. Это обеспечивает поддержание ионного и молекулярного состава клетки.
4. Мембрана не имеет свободных концов. Она всегда замыкается в пузырьки.
5. Асимметричность. Состав наружного и внутреннего слоев как белков, так и липидов различен.
6. Полярность. Внешняя сторона мембраны несёт положительный заряд, а внутренняя – отрицательный.
ФУНКЦИИ МЕМБРАНЫ
1) Барьерная – плазмалемма отграничивает цитоплазму и ядро от внешней среды. Кроме того, мембрана делит внутреннее содержимое клетки на отсеки (компартменты), в которых зачастую протекают противоположные биохимические реакции.
2) Рецепторная (сигнальная) – благодаря важному свойству белковых молекул – денатурации, мембрана способна улавливать различные изменения в окружающей среде. Так, при воздействии на мембрану клетки различных средовых факторов (физических, химических, биологических) белки, входящие в ее состав, меняют свою пространственную конфигурацию, что служит своеобразным сигналом для клетки. Это обеспечивает связь с внешней средой, распознавание клеток и их ориентацию при формировании тканей и т.д. С этой функцией связана деятельность различных регуляторных систем и формирование иммунного ответа.
3) Обменная – в состав мембраны входят не только структурные белки, которые образуют ее, но и ферментативные, являющиеся биологическими катализаторами. Они располагаются на мембране в виде «каталитического конвейера» и определяют интенсивность и направленность реакций метаболизма.
9 вопрос как продолжение 8.
4) Транспортная – молекулы веществ, диаметр которых не превышает 50 нм, могут проникать путем пассивного и активного транспорта через поры в структуре мембраны. Крупные вещества попадают в клетку путем эндоцитоза (транспорт в мембранной упаковке), требующего затраты энергии. Его разновидностями являются фаго- и пиноцитоз.
Пассивный транспорт – вид транспорта, в котором перенос веществ осуществляется по градиенту химической или электрохимической концентрации без затраты энергии АТФ. Выделяют два вида пассивного транспорта: простая и облегченная диффузия. Диффузия – это перенос ионов или молекул из зоны более высокой их концентрации в зону более низкой концентрации, т.е. по градиенту.
Простая диффузия – ионы солей и вода проникают через трансмембранные белки или жирорастворимые вещества по градиенту концентрации.
Облегченная диффузия – специфические белки-переносчики связывают вещество и переносят его через мембрану по принципу «пинг-понга». Таким способом через мембрану проходят сахара и аминокислоты. Скорость такого транспорта значительно выше, чем простой диффузии. Кроме белков- переносчиков, в облегченной диффузии принимают участие некоторые антибиотики – например, грамитидин и ваномицин. Поскольку они обеспечивают транспорт ионов, их называют ионофорами.
Активный транспорт – это вид транспорта, при котором расходуется энергия АТФ, он идёт против градиента концентрации. В нем принимают участие ферменты АТФ-азы. В наружной клеточной мембране находятся АТФ-азы, которые осуществляют перенос ионов против градиента концентрации, это явление называется ионным насосом. Примером является натрий-калиевый насос. В норме в клетке больше ионов калия, во внешней среде – ионов натрия. Поэтому по законам простой диффузии калий стремится из клетки, а натрий – в клетку. В противовес этому натрий-калиевый насос накачивает против градиента концентрации в клетку ионы калия, а ионы натрия выносит во внешнюю среду. Это позволяет поддерживать постоянство ионного состава в клетке и её жизнеспособность. В животной клетке одна треть АТФ расходуется на работу натрий-калиевого насоса.
Разновидностью активного транспорта является транспорт в мембранной упаковке – эндоцитоз. Крупные молекулы биополимеров не могут проникать через мембрану, они поступают в клетку в мембранной упаковке. Различают фагоцитоз и пиноцитоз. Фагоцитоз – захват клеткой твердых частиц, пиноцитоз – жидких частиц. В этих процессах выделяют стадии:
1) узнавание рецепторами мембраны вещества; 2) впячивание (инвагинация) мембраны с образованием везикулы (пузырька); 3) отрыв пузырька от мембраны, слияние его с первичной лизосомой и восстановление целостности мембраны; 4) выделение непереваренного материала из клетки (экзоцитоз).
Эндоцитоз является способом питания для простейших. У млекопитающих и человека имеется ретикуло-гистио-эндотелиальная система клеток, способная к эндоцитозу – это лейкоциты, макрофаги, клетки Купфера в печени.
ОСМОТИЧЕСКИЕ СВОЙСТВА КЛЕТКИ
Осмос – односторонний процесс проникновения воды через полупроницаемую мембрану из области с меньшей концентрацией раствора в область с более высокой концентрацией. Осмос обусловливает осмотическое давление.
Диализ – односторонняя диффузия растворенных веществ.
Раствор, в котором осмотическое давление такое же, как и в клетках, называют изотоническим. При погружении клетки в изотонический раствор её объем не изменяется. Изотонический раствор называют физиологическим – это 0,9% раствор хлорида натрия, который широко применяется в медицине при сильном обезвоживании и потери плазмы крови.
Раствор, осмотическое давление которого выше, чем в клетках, называют гипертоническим. Клетки в гипертоническом растворе теряют воду и сморщиваются. Гипертонические растворы широко применяются в медицине. Марлевая повязка, смоченная в гипертоническом растворе, хорошо впитывает гной.
Раствор, где концентрация солей ниже, чем в клетке, называют гипотоническим. При погружении клетки в такой раствор вода устремляется в нее. Клетка набухает, ее тургор увеличивается, и она может разрушиться. Гемолиз – разрушение клеток крови в гипотоническом растворе.
Осмотическое давление в организме человека в целом регулируется системой органов выделения.
5 вопрос.
ЦИТОПЛАЗМА
Цитоплазма (греч. citos – клетка, plazma – вылепленная) – это внутренняя среда клетки. Включает гиалоплазму, цитоскелет, органоиды и включения.
❇ Гиалоплазма (матрикс) заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Это тонкозернистое, полупрозрачное, вязкое, студенистое вещество цитоплазмы.
Химический состав. Гиалоплазма – это коллоидный раствор с высоким содержанием воды и белков. Гиалоплазма способна переходить из золеобразного (жидкого) состояния в гелеобразное. Состав гиалоплазмы определяет осмотические свойства клетки.
Н2О 70 – 75%,
белки 10 – 20%,
липиды 1 – 5%,
углеводы 0,2 – 2%,
нуклеиновые кислоты 1 – 2%,
минеральные соединения 1 – 1,5%,
АТФ и другие низкомолекулярные органические вещества 0,1 – 0,5%.
Функции: 1) транспортная: обеспечивает перемещение веществ в клетке;
2) обменная: является средой для протекания химических реакций внутри клетки;
3) собственно внутренняя среда клетки, в которую погружены все другие компоненты цитоплазмы и ядро.
❇ Органоиды – это постоянные структуры цитоплазмы, выполняющие в клетке определенные функции. Исходя из мембранного принципа строения и функциональной принадлежности, все органоиды клетки делятся на две большие группы: органоиды общего и специального назначения.
Органоиды специального значения присутствуют у простейших (органоиды движения – ложноножки, реснички, жгутики ), органоид осморегуляции – сократительная вакуоль, органоиды защиты и нападения – трихоцисты, светочувствительный глазок – стигма) и в специализированных клетках многоклеточных организмов (реснички, жгутики, микроворсинки).
Органоиды общего значения встречаются абсолютно во всех эукариотических клетках и подразделяются на немембранные и мембранные.
К немембранным органоидам клетки общего значения относятся рибосомы, клеточный центр (центросома), микротрубочки, микрофиламенты и промежуточные филаменты (микрофибриллы).
Мембранные органоиды могут быть одно- и двумембранные.
Одномембранный принцип строения имеют эндоплазматическая сеть (ЭПС), комплекс Гольджи, лизосомы, пероксисомы и растительные вакуоли. Одномембранные органоиды клетки объединяются в вакуолярную систему, компоненты которой представляют собой отдельные или связанные друг с другом отсеки, распределенные закономерным образом в гиалоплазме. Так, различные вакуоли (вакуоли растительных клеток, пероксисомы, сферосомы и др.) возникают из пузырьков эндоплазматического ретикулума, в то время как лизосомы – из пузырьков вакуолярного комплекса аппарата Гольджи.
Двумембранными органоидами клетки являются митохондрии и пластиды (лейкопласты, хлоропласты и хромопласты).
10 вопрос.
Жизненный цикл клеток
В настоящее время считается бесспорным, что клетки возникают только в результате деления. При этом вновь возникшие клетки не сразу имеют все структурные компоненты, свойственные зрелым клеткам. Для созревания клеток должно пройти определенное время. Процесс созревания клеток называется дифференцировкой. В процессе дифференцировки клетки приобретают черты структурной и функциональной зрелости. Большинство клеток по истечении определенного времени приступает к делению (за исключением зрелых нейронов) или же погибает. Генетически запрограммированная гибель клеток называется апоптозом. В отличие от последней, гибель клеток в результате случайных внешних причин, например, травмы, ожога и т.д., называется некрозом.
Деление клеток лежит в основе роста организма и регенерации. Основным способом деления соматических клеток является митоз. Кроме него, встречаются и другие формы деления: амитоз и эндомитоз.
Промежуток времени между клеточными делениями называется интерфазой.
Некоторые цитологи выделяют два вида интерфаз: гетеросинтетическую и аутосинтетическую.
В период гетеросинтетеической интерфазы клетки работают на организм, выполняя свои функции составного компонента того или иного органа или такни. В период аутосинтетической интерфазы клетки готовятся к митозу или мейозу. В этой интерфазе выделяют три периода: пресинтетический – G1, синтетический – S, и постсинтетический – G2.
Период G1 – самая продолжительная фаза. В этот период клетка синтезирует РНК и белки.
В S-периоде продолжается синтез белка и происходит репликация ДНК. В большинстве клеток этот период длится 8-12 часов.
В G2 – периоде продолжается синтез РНК и белка (например, тубулина для построения микротрубочек веретена деления). Происходит накопление АТФ для энергетического обеспечения последующего митоза. Эта фаза длится 2-4- часа.
Кроме интерфазы, для характеристики временной организации клеток выделяют такие понятия, как жизненный цикл клеток, клеточный цикл и митотический цикл. Под жизненным циклом клетки понимают время жизни клетки с момента ее возникновения после деления материнской клетки и до конца ее собственного деления или же до гибели.
Клеточный цикл – это совокупность процессов, протекающих в аутосинтетическую интерфазу, и собственно митоз.
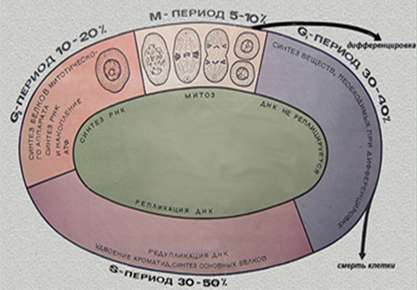
Рис.27. Клеточный цикл. Интерфаза.
Митотический цикл – это совокупность процессов, протекающих в клетке в течение собственно митоза.
Вопрос. МИТОЗ
Митоз (от греч. митос – нить), или кариокинез (греч. карион – ядро, кинезис – движение), или непрямое деление. Это процесс, в ходе которого происходит конденсация хромосом и равномерное распределение дочерних хромосом между дочерними клетками. Митоз включает в себя пять фаз: профаза, прометафаза, метафаза, анафаза и телофаза. В профазе хромосомы конденсируются (скручиваются), становятся заметными и располагаются в виде клубка. Центриоли делятся на две и начинают двигаться к клеточным полюсам. Между центриолями появляются нити, состоящие из белка тубулина. Происходит образование митотического веретена. В прометафазе ядерная оболочка распадается на мелкие фрагменты, а погруженные в цитоплазму хромосомы начинают двигаться к экватору клетки. В метафазе хромосомы устанавливаются на экваторе веретена и становятся максимально компактизированными. Каждая хромосома состоит из двух хроматид, связанных друг с другом центромерами, а концы хроматид расходятся, и хромосомы принимают Х-образную форму. В анафазе дочерние хромосомы (бывшие сестринские хроматиды) расходятся к противоположным полюсам. Предположение о том, что это обеспечивается сокращением нитей веретена, не подтвердилось.

Рис.28. Характеристика митоза и мейоза.
Многие исследователи поддерживают гипотезу скользящих нитей, согласно которой соседние микротрубочки веретена деления, взаимодействуя друг с другом и сократительными белками, тянут хромосомы к полюсам. В телофазе дочерние хромосомы достигают полюсов, деспирализуются, образуется ядерная оболочка, восстанавливается интерфазная структура ядер. Затем наступает разделение цитоплазмы – цитокинез. В животных клетках этот процесс проявляется в перетяжке цитоплазмы за счет втягивания плазмолеммы между двумя дочерними ядрами, а в растительных клетках мелкие пузырьки ЭПС, сливаясь, образуют изнутри цитоплазмы клеточную мембрану. Целлюлозная клеточная стенка образуется за счет секрета, накапливающегося в диктиосомах.
Продолжительность каждой из фаз митоза различна – от нескольких минут до сотен часов, что зависит как от внешних, так и внутренних факторов и типа тканей.
Нарушение цитотомии приводит к образованию многоядерных клеток. При нарушении репродукции центриолей могут возникнуть многополюсные митозы.
АМИТОЗ
Это прямое деление ядра клетки, сохраняющего интерфазную структуру. При этом хромосомы не выявляются, не происходит образования веретена деления и их равномерного распределения. Ядро делится путем перетяжки на относительно равные части. Цитоплазма может делиться перетяжкой, и тогда образуются две дочерние клетки, но может и не делиться, и тогда образуются двуядерные или многоядерные клетки.

Рис.29. Амитоз.
Амитоз как способ деления клеток может встречаться в дифференцированных тканях, например, скелетных мышцах, клетках кожи, а также в патологических изменениях тканях. Однако он никогда не встречается в клетках, нуждающихся в сохранении полноценной генетической информации.
ЭНДОМИТОЗ
Эндомитоз – процесс, который приводит к образованию полиплоидных клеток. Это происходит вследствие блокады митоза либо сразу после премитотического периода интерфазы(G2), либо на стадии профазы или метафазы, после чего клетки переходят в состояние интерфазы и могут приступить опять к репликации ДНК, что приводит к полиплоидии.
 Рис.30. Эндомитоз.
Рис.30. Эндомитоз.
Появление полиплоидных соматических клеток может происходить и в результате отсутствия цитотомии. Если такая двуядерная клетка снова вступит в синтетический период интерфазы, то оба ядра становятся не диплоидными, а тетраплоидными. Подобным образом образуются полиплоидные клетки в печени, эпителии мочевого пузыря и др. Характерно, что полиплоидизация встречается на терминальных этапах развития клеток, как правило, характерна для специализированных, дифференцированных клеток и не встречается при образовании половых и стволовых клеток, а также в ходе эмбриогенеза (исключая провизорные органы).
Регуляция клеточного деления. Целостность тканей и органов может поддерживаться только в том случае, если рост и деление каждой индивидуальной клетки будут запрограммированы и скоординированы с делением соседних клеток. У млекопитающих рост и деление клеток контролируются различными внешними факторами по принципу обратной связи. К таким факторам относятся: наличие свободного пространства, на котором могут распластываться клетки, секреция окружающими клетками стимулирующих или ингибирующих веществ. Существует модель регуляции запуска митоза с помощью какого-то нестабильного, триггерного белка. При этом такие неблагоприятные факторы, как увеличение плотности клеток или клеточное голодание, снижают синтез белка и останавливают клеточный цикл на периоде G1 интерфазы. Обсуждаются и другие факторы.
Вопрос.
МЕЙОЗ
Мейоз (греч. мейозис – уменьшение) имеет место на стадии созревания гамет. Благодаря мейозу из диплоидных незрелых половых клеток образуются гаплоидные гаметы: яйцеклетки и сперматозоиды. Мейоз включает в себя два деления: редукционное (уменьшительное) и эквационное (уравнительное), каждое из которых имеет те же фазы, что и митоз. Однако, несмотря на то, что клетки делятся два раза, удвоение наследственного материала происходит только один раз – перед редукционным делением - и отсутствует перед эквационным.
Цитогенетический результат мейоза (образование гаплоидных клеток и перекомбинация наследственного материала) происходит во время первого (редукционного) деления. Оно включает 4 фазы: профазу, метафазу, анафазу и телофазу.
Профаза I подразделяется на 5 стадий: лептонемы (лептотены), или стадия тонких нитей, зигонемы (зиготены), стадия пахинемы (пахитены), или толстых нитей, стадия диплонемы (диплотены) и стадия диакинеза.

Рис.31. Мейоз. Процессы, происходящие при редукционном делении.
В стадии лептонемы происходит спирализация хромосом и их выявление в виде тонких нитей с утолщениями по длине. В стадии зигонемы продолжается компактизация хромосом, а гомологичные хромосомы сближаются попарно и конъюгируют: каждая точка одной хромосомы совмещается с соответствующей точкой гомологичной хромосомы (синапсис). Две рядом лежащие хромосомы образуют биваленты.
В пахинеме между хромосомами, составляющими бивалент, может происходить обмен гомологичными участками (кроссинговер). На этой стадии видно, что каждая конъюгирующая хромосома состоит из двух хроматид, а каждый бивалент – из четырех хроматид (тетрад).
Диплонема характеризуется, появлением сил отталкивания конъюгатов начиная от центромер, а затем и в других участках. Хромосомы остаются связанными между собой только в местах кроссинговера.
В стадии диакинеза (расхождение двойных нитей) парные хромосомы частично расходятся. Начинается формирование веретена деления.
В метафазе I пары хромосом (биваленты) выстраиваются по экватору веретена деления, образуя метафазную пластинку.
В анафазе I к полюсам расходятся двухроматидные гомологичные хромосомы, и на клеточных полюсах скапливается их гаплоидный набор. В телофазе 1 происходят цитотомия и восстановление структуры интерфазных ядер, каждое из которых содержит гаплоидное число хромосом, но диплоидное количество ДНК (1n2c). После редукционного деления клетки переходят в короткую интерфазу, во время которой не наступает период S, и начинается эквационное (2-е) деление. Оно протекает, как обычный митоз, в результате чего образуются половые клетки, содержащие гаплоидный набор однохроматидных хромосом (1n1c)

Рис.32. Мейоз. Эквационное деление.
Таким образом, во время второго мейотического деления количество ДНК приводится в соответствие с количеством хромосом.
13 вопрос.
Гаметогенез – это последовательный процесс, который обеспечивает размножение, рост и созревание половых клеток в мужском организме (сперматогенез) и женском (овогенез).
Гаметогенез протекает в половых железах - сперматогенез в семенниках у мужчин, а овогенез в яичниках у женщин. В результате гаметогенеза в организме женщины образуются женские половые клетки - яйцеклетки, а у мужчин - мужские половые клетки сперматозоиды.
Именно процесс гаметогенез (сперматогенез, овогенез) дает возможность мужчине и женщине возможность воспроизведения потомства.
Стадии овогенеза и сперматогенеза
Гаметогенез имеет несколько стадий. Сходство сперматогенеза и овогенеза как раз и заключается в том, что три стадии у них одинаковы.
1. Стадия размножения. Первичные клетки на этой стадии называются сперматогониями и овогониями, из них в последующем образуются мужские и женские половые клетки. Половые клетки несколько раз делятся путем митоза, и количество их значительно возрастает. Сперматогонии размножаются у мужчины в течение всего репродуктивного периода, а размножение овогоний происходит в эмбриональном периоде и наиболее интенсивно происходит во 2 - 5 месяц внутриутробного развития.
2. Стадия роста. В этот период клетки значительно увеличиваются в размерах. Сперматогонии и овогонии превращаются в сперматоциты и овоциты I порядка. Овоциты I порядка достигают больших размеров, поскольку накапливают питательные вещества.
3. Стадия созревания. На этой стадии происходят два следующих друг за другом деления - мейоз I и мейоз II. После первого деления образуются сперматоциты и овоциты II порядка, а после второго деления - сперматиды и зрелые яйцеклетки с тремя полярными тельцами, которые в процессе размножения не участвуют и погибают. При созревании один сперматоцит I порядка дает четыре сперматиды, а один овоцит I порядка образует одну яйцеклетку и три полярных тельца.
Эти особенности сперматогенеза и овогенеза имеют биологический смысл, который связан с разным назначением мужских и женских гамет. Неравномерное деление клеток при овогенезе (меньше) обеспечивает формирование крупной яйцеклетки, в ней накапливается большее количество питательных веществ, так как из оплодотворенного яйца будет развиваться новый организм.
При сравнительной характеристике овогенеза и сперматогенеза можно заметить, что сперматозоидов образуется значительно больше, и это также имеет биологический смысл.
Яйцеклетку достигает только один сперматозоид, проникает в нее и доставляет свой набор хромосом. Остальные же в процессе поиска яйцеклетки массово погибают.
При сравнении овогенеза и сперматогенеза становится понятным, почему сперматозоидам нет необходимости в запасании питательных веществ - их существование кратковременно, а подвижность должна быть высокой.
 4. Стадия формирования. Она характерна только для сперматогенеза. Незрелая сперматида превращается в сперматозоид, приобретая свойственный ему вид. Образование сперматозоидов у мужчин начинае
4. Стадия формирования. Она характерна только для сперматогенеза. Незрелая сперматида превращается в сперматозоид, приобретая свойственный ему вид. Образование сперматозоидов у мужчин начинае