Левая колонка - усреднение первых 49 проб теста, правая - последних 49 проб. Бин
- 256 мс. СС S - мозолистое тело левого полушария. Остальные обозначения - как на рис. 18 и 21
Первая позиция вновь подтверждает положение о том, что обеспечение мыслительной деятельности осуществляется мозговой системой, а не мозгом «в целом». Вторая - наглядно демонстрирует разницу мозговых систем обеспечения функций, наличие в мозговой системе обеспечения, например, двигательной деятельности (в данном случае - речедвигательной), значительно большего числа жестких элементов. Таким образом, в исследованиях нейронных коррелятов мыслительных процессов было подтверждено высказанное ранее предположение о связи фактора гибкости в мозговых системах со сложностью (нестереотипностью) обеспечиваемой деятельности (Бехтерева, 1971, 19806).
Данный этап исследований по отношению к первому может быть обозначен не как второй этап, а точнее, как второй виток спирали в изучении проблемы, так как, по существу, основная проблема сохранялась, однако основной задачей оказывалось обнаружение уже не просто коррелятов мыслительных процессов, но, как указывалось, - статистически достоверных коррелятов. Исследования были прежде всего сосредоточены на изучении перестроек частоты импульсной активности, однако рассматривались с тех же методологических позиций и перестройки структуры динамики импульсной активности. Проводились поиски и изучение не только конкретных последовательностей интервалов, но более общей связи (и прежде всего, ее наличия) между реализацией мыслительной деятельности и изменениями структуры импульсной активности нейронных популяций. Использовался показатель нормированной гистограммы плотности распределения межимпульсных интервалов, то есть принципиально - отношения исследуемых интервалов к средней величине межепайкового интервала. A priori можно было предполагать, что если в этих условиях и выявятся какие-то изменения, то они явятся производным перестройки частоты импульсного потока. Однако, как показали работы Н. Г. Шкуриной (1984), наблюдался не этот один, а три (все три!) теоретически возможных варианта соотношений частоты и структуры импульсной активности. Регистрировались достоверные изменения частоты без достоверных изменений структуры; наблюдалось сочетанное изменение обоих показателей; и наконец, что особенно интересно, изменения структуры потока, как и в предыдущих двух ситуациях, соотносимые с фазами психологического теста, могли наблюдаться изолированно, без существенного увеличения частоты импульсной активности (рис. 23, а, б, в).
Анализировалась импульсная активность 5 популяций вентролатерального ядра зрительного бугра, 2 - бледного шара, 3 - хвостатого ядра и активность, отводимая из мозолистого тела. Следует отметить, что различные соотношения динамики частоты импульсной активности могли наблюдаться как в разных популяциях и, таким образом, в зависимости от зоны мозга, так и в одной и той же популяции, но уже в зависимости от характера теста. Эти материалы подтвердили на новом уровне, что изменения структуры импульсного потока были правомерно отнесены к нейрофизиологическим коррелятам мыслительной деятельности. Эти же данные показали, что перестройки структуры импульсной активности могут иметь в этом плане самостоятельное значение, а отсюда допустимо рассматривать перестройки частоты и структуры импульсной активности как имеющие взаимодополняющее значение. Доказательства значения перестроек структуры импульсной активности как одного из нейрофизиологических коррелятов мыслительной деятельности были получены также в исследованиях С. В. Медведева и Н. И. Цыганкова (1985), проведенных способом авторегрессионного анализа (рис. 24, 25).

Рис. 23.
Нормированные гистограммы плотности распределения межимпульсных интервалов и ПСГ активности нейронной популяции в бледном шаре правого полушария при выполнении тестов на опознание (а, б) и запоминание (в).
Вверху - нормированные гистограммы распределения межимпульсных интервалов, соответствующие выбранным эпохам анализа (ограничены вертикальными штриховыми линиями) (по оси абсцисс - величина межимпульсного интервала, ед. стандартного отклонения - т; по оси ординат - число межимпульсных интервалов, имеющих данную величину); внизу - ПСГ (по оси абсцисс - время: бин - 150 мс; по оси ординат - число импульсов в бине, усредненное по заданному числу проб: а - 53 пробы, б - 53, в - 107). Заштрихована область высокодостоверного ответа на ПСГ. Стрелками указаны моменты предъявления стимулов и начала ответов испытуемого. Точками отмечены гистограммы, значимо отличающиеся по форме от гистограмм, накопленных по двум другим эпохам анализа
С помощью приема линейной авторегрессионной функции 4-го порядка (вычислялись первые 4 коэффициента) исследовались нейронные популяции, расположенные в стриопаллидарной системе, лимбических образованиях и различных

Рис. 25. Зависимость нормированного структурного коэффициента Y -от уровня амплитудной дискриминации в двух исследованиях (а, б) у одного больного.
По оси абсцисс - среднее значение текущей частоты при различных уровнях

Рис. 24. Динамика
структурных коэффициентов b -во время выполнения корректурного теста (мозолистое тело). Вертикальными линиями разделены последовательные повторения задания. Штриховой линией показан уровень b =0.1. Как видно, импульсная активность структурирована на уровне b. Динамика b -отражает временную структуру теста
областях коры. Прием позволил выделить из популяций такие субпопуляции, в которых импульсная активность была структурирована. При этом обнаружены своего рода сверхстабильные ансамбли, зависимые от условий исследования. Этот способ анализа оказался выгодным для выделения из нейронной популяции ансамбля, группы нейронов, образующих рабочую единицу при выполнении какой-либо деятельности.
амплитудной дискриминации, Гц. Штриховыми линиями ограничен уровень достоверности Р = 0.05. Видна зависимость структуры потока от уровня дискриминации, то есть от размера регистрируемой субпопуляции. Обозначения структур мозга - как на рис. 18 и 21
В исследовании мозговых систем обеспечения деятельности безусловно необходимо изучение местных событий в различных зонах мозга и динамики этих местных событий. На этой основе появляются известные основания для суждения о свойствах мозговой системы обеспечения деятельности. Однако исследование собственно системы должно включать и исследование взаимоотношений между активными зонами мозга - предположительно - звеньями системы.
Такого рода исследования проводились приемом корреляционного анализа, с изучением соотношений в разных зонах мозга изменений частоты импульсной активности и методом анализа динамики совпадений разрядов в микроинтервалы времени в различных зонах мозга.
Исследование соотношений частоты разрядов в различных зонах мозга, активных при осуществлении психологических тестов наряду с самим фактом наличия корреляции между этими зонами показало отсутствие обязательной зависимости тесноты связи между различными зонами мозга от местных изменений частоты импульсной активности (рис. 26). Это позволило рассматривать и третий показатель - соотношение активности в звеньях мозговых систем- также как дополнительный мозговой механизм в обеспечении мыслительных процессов. Таким образом, нами рассматриваются как взаимодополняющие механизмы в нейрофизиологическом кодировании мыслительных процессов изменения частоты, структуры импульсной активности и взаимоотношений этой активности в звеньях мозговой системы.
Как видно из приведенного рисунка (см. рис. 26), по-видимому, следует в прямое развитие сказанного подчеркнуть, что уменьшение выраженности изменений частоты импульсной активности, вплоть до исчезновения достоверных сдвигов, еще не может расцениваться как выключение зон из обеспечения деятельности. И наоборот, простой прием корреляционного анализа может использоваться для уточнения степени: действительно ли зона мозга перестала участвовать в обеспечении деятельности как звено системы.

Рис. 26. ПСГ
импульсной активности, зарегистрированной в лучистом венце (левая колонка), премоторной коре правого полушария (правая колонка), и динамика коэффициента корреляции между текущей частотой разрядов этих популяций на протяжении пробы (средняя колонка).
Верхний ряд гистограмм - данные для проб первой половины теста на сравнение по величине двух цифр (49 проб); нижний ряд - данные для проб второй половины теста (48 проб). F - частота, F - средняя частота в бине, R - коэффициент корреляции. Остальные обозначения - как на рис. 18
Исследование динамики совпадений разрядов в зонах мозга в микроинтервалы времени проводилось по принципиальной схеме, предложенной Герштейном (Gerstein et al., 1978) и адаптированной Е. М. Кроль и С. В. Медведевым (1982).
Осуществлялось исследование достоверного увеличения числа совпадений импульсной активности в различных популяциях, статистический анализ неслучайного числа совпадений в мозговых зонах, расположенных в области мозговых образований.
Основным психологическим тестом, используемым на данном этапе для исследования соотношений импульсной активности в различных зонах мозга, был так называемый корректурный тест. Перед больным на экране проецировалась таблица из 14 строчек колец с разрывами, имеющими 8 различных ориентации. В некоторые строчки были включены и полностью замкнутые кольца. Предлагалось считать кольца с определенной ориентацией разрыва или отыскивать замкнутое кольцо, которое, кстати, не обязательно присутствовало на слайде. Каждый вид пробы повторялся по 8 раз. Слайд предъявлялся несколько быстрее, чем можно было полностью просмотреть все строки таблицы (около 150 с). Такого рода монотонный длительный тест использовался с целью активации и дальнейшего поддержания в мозгу в активном состоянии определенной системы. Предполагалось, что в этих условиях взаимодействия между звеньями этой системы могут быть изучены наилучшим образом. Исследования С. В. Медведева и сотрудников (Бехтерева и др., 1984; Медведев, Белов, 1986) подтвердили эффективность данной методики и показали, что количество совпадений импульсных разрядов нейронов между дистантными звеньями системы в большей мере определялось характером выполняемой деятельности (рис. 27, 28). Было показано также, что существенное увеличение или, наоборот, уменьшение числа совпадений разрядов по отношению к фоновому уровню может наблюдаться в процессе реализации пробы между некоторыми мозговыми зонами в очень короткие интервалы времени, в том числе в интервалы длительностью 1-2 мс. Эти факты поставили задачу специального рассмотрения физиологических условий такого рода синхронизации разрядов, так как в этом случае трудно было понять эту синхронизацию без введения представлений о пейсмекерном механизме (см. выше представления о динамическом пейсмекере) или о возможности скоростного взаимодействия между дистантными зонами мозга.

Рис. 27. Динамика
связанности между дистантно расположенными нейронными популяциями. I - нейронные участки мозговых структур; II - фон (отсутствие целенаправленной деятельности); III - корректурные пробы со счетом; IV - корректурные пробы со счетом, перемежающиеся сюжетом; V - корректурные пробы без счета. По оси
абсцисс штрихами обозначены последовательные пробы; по оси ординат отложена величина квантили (для Р < 0.975). Калибровка по вертикали (длина штриха в колонке IV); квантиль равна 2.0. Сплошные вертикальные линии - корректурные пробы, штриховые - сюжетные пробы. Характеристики связей: T=1 с, т=1 мс

Рис. 28. Паттерны стабильной связанности нейронных популяций.
А—Д - больные, черными точками указаны исследованные популяции. Жирными линиями соединены стабильно связанные популяции
Эту последнюю возможность нельзя исключить полностью в связи с обнаруженным
в специальных условиях наблюдения очень резким падением импеданса между двумя звеньями системы, в этих специальных условиях связанной с обеспечением движения (Бехтерева и др., 1980). В связи с трудностью исследования этого вопроса у человека, вероятно, следует организовать целенаправленное его изучение в эксперименте на животных, тем более что в данной ситуации совершенно не обязательно проводить исследования именно системы обеспечения мыслительных процессов.
В проведенных работах достаточно неожиданно для тех, кто конструировал тесты, было обнаружено, что длительное осуществление однообразной деятельности совсем не обязательно коррелирует с однообразием ситуация в мозгу. Было показано, что соотношения в системе обеспечения мыслительной деятельности перестраиваются независимо от длительности монотонного теста на протяжении 0.5-1 с. Иными словами, повышенная синхронизация разрядов нейронов, улавливаемая методом совпадений, наблюдается попеременно между различными активными зонами мозга. Высокая синхронизация активности нейронов в каждый данный микроинтервал времени может наблюдаться далеко не между всеми активными точками, что может быть проиллюстрировано рис. 29.
Следует подчеркнуть не только динамичность связей, но и возможность как бы независимых связей между парами нейронных популяций. Нетранзитивность связей была обнаружена независимо от расстояния между исследованными зонами. Кроме того, по аналогии с местными явлениями были обнаружены и связи разной степени жесткости. Таким образом, и в этом случае была обнаружена динамичность мозгового обеспечения мыслительной деятельности, в данном случае в форме динамической реорганизации системы на протяжении монотонной деятельности. Если в научном исследовании позволены образные выражения (а почему бы и нет?), создается впечатление, что мозг как бы «борется» против монотонности. Динамичность, которую мы наблюдали в местных процессах, является, по-видимому, важным общим принципом работы мозга.
Рис. 29. Схематическое представление элементов системы обеспечения монотонной деятельности (корректуры) с сохранением функциональной структуры действия.
Вверху - корректурные пробы со счетом, внизу - без счета. Характеристики связей: Т=1с, т=1мс
Что же касается самого методического приема - исследования совпадений разрядов, то он оказался адекватным именно для изучения соотношений активности в небольшие интервалы времени, давал своего рода «мгновенные снимки» ситуации в системе обеспечения мыслительных процессов. При больших бинах (больше, чем 1-3 мс) перестройки, связанные с заданной деятельностью, не выявлялись (Бехтерева и др., 1984). Отсюда, по-видимому, для исследования взаимоотношения местных процессов в звеньях мозговых систем (активных при какой-то деятельности зонах мозга) целесообразно использовать не только один, но целый ряд приемов, в том числе и таких, которые не только позволяют выявить соотношение активности в различных зонах мозга, но и вскрыть различные механизмы организации системы.
Третий этап исследований может быть характеризован как попытка минимизации дефектов методов анализа, связанных с накоплением и таким образом усреднением данных, то есть как бы вновь, на новом витке спирали, попытаться использовать для изучения мозга приемы, более адекватные механизмам мозга. В этом плане предложен и используется компонентный анализ (Гоголицын, Пахомов, 1984, 1985), резервы которого, повидимому, еще не полностью раскрыты. Принцип метода - в первоначальном сохранении максимума информации о динамике местных перестроек в мозгу. Проводится количественное описание импульсной активности в рамках более гибких представлений о паттернах текущей частоты. Это достигается представлением физиологических мозговых событий по каждой пробе в виде диаграмм таким образом, что по оси абсцисс отложено время развития перестроек частоты (ее учащения или
урежения), а по оси ординат - выраженность этих перестроек в форме амплитудного показателя (количество импульсов в бине).
Последовательное автоматизированное наложение на одно изображение (диаграмму) данных по множеству проб далее сопровождается выделением кластеров, то есть выделением областей максимумов сгущений точек (рис. 30). Сравнение получаемых с помощью данного приема результатов с ПСГ показало, что ПСГ, во- первых, действительно может отражать события, достаточно близкие к тому, что происходит в импульсной активности нейронов по ходу всего теста, во-вторых, может отражать их приблизительно и, наконец, за низкоамплитудной ПСГ могут скрываться существенные отклонения импульсной активности от фона, но различные по направленности, амплитуде и времени развития на разных фазах теста в каждой пробе (рис. 31). Применение компонентного анализа позволило, таким образом, продемонстрировать, из каких реальных физиологических событий, происходящих при однократных воздействиях стимула, складывается усредненная картина ПСГ.


Рис. 30. Схематическое
изображение основных этапов компонентного анализа импульсной активности нейронов.
Импульсная активность регистрируется при выполнении психологических тестов, состоящих из однотипных проб. Исходным материалом для анализа являются кривые, описывающие изменения частоты разрядов нейронов в отдельных пробах. Каждая такая кривая с помощью специального алгоритма раскладывается на компоненты, то есть представляется в виде суммы колоколообразных пиков, характеризуемых латентностью, амплитудой и длительностью. Это позволяет получить компонентные представления реакций в отдельных пробах. Используя латентные периоды и амплитуды всех компонентов, можно построить диаграмму рассеяния, наглядно иллюстрирующую степень вариабельности реакций в отдельных пробах на протяжении теста.
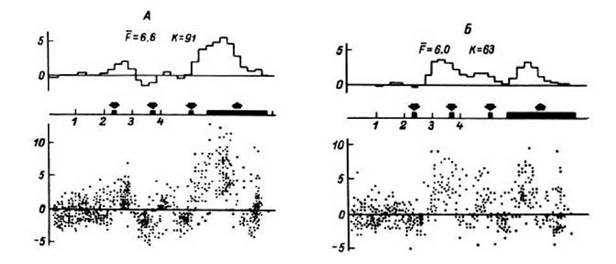

Рис. 31. ПСГ и диаграммы рассеяния «латентный период—амплитуда» активности нейронных популяций в хвостатом ядре левого полушария (А), мозолистом теле левого полушария (Б) и премоторной коре правого полушария (В) в тесте на сравнение цифр по величине.
Под ПСГ на оси времени (цена деления - 1 с) черными прямоугольниками отмечены эпохи предъявления (стрелки вниз) символов (цифра, буква, цифра) и звучание ответа испытуемого (стрелка вверх). По оси ординат ПСГ - отклонение частоты разрядов (имп./бин) от среднего уровня в фоне. Бин ПСГ - 256 мс. F - средний уровень частоты разрядов (имп./бин) на протяжении фонового фрагмента (от начала пробы до момента предъявления первой цифры). К - число проб. ПСГ и диаграмма рассеяния построены в одинаковом масштабе.
Вторым важным направлением данного этапа исследований является сравнительная характеристика функциональных свойств различных мозговых зон. С этой целью анализ динамики импульсной активности ряда нейронных популяций подкорки и коры (чаще всего способом ПСГ) осуществляется при предъявлении больному не какого-то одного, а совокупности, батареи, тестов. В этом случае в полном соответствии с более ранними данными о полифункциональности нейронных популяций (Бехтерева, 1971) были обнаружены разные реакции импульсной активности нейронов коры и подкорковых зон мозга на психологические тесты с различием конкретныхзаданий в пробах (Кропотов, Пономарев, 1985). Так, нарис. 31 представлены ПСГ, построенные на основе анализа импульсной активности нейронных популяций, расположенных в различных зонах коры и подкорковых структур. Как видно из рис. 32, ПСГ могли различаться при разных психологических пробах очень значительно. На данном рисунке наибольшая изменчивость в зависимости от теста наблюдалась в отведениях одной из популяций бледного шара, вентролатерального ядра зрительного бугра и теменной коры (в области поля 40, по Бродману).
и
Нажимает кнопку 0.5 с

Рис. 32. Примеры ПСГ, вычисленных для различных нейронных популяций корковых и подкорковых структур, в тесте с предъявлением зрительных стимулов, вызывающих различные ответные реакции больных.
S - предъявление стимула, определяющего ответную реакцию; S - предъявление паттерна шахматной доски, служащего сигналом для ответной реакции. Реакции больного в ответ на предъявление стимулов следующие: буква П - игнорируется, буква Е - подсчитывается текущее число таких стимулов, цифры называются вслух, буква Н - больной нажимает кнопку. Стрелками указаны структуры, в которых регистрировалась импульсная активность. Зачернены участки ПСГ, соответствующие уровню значимости Р < 0.01
Аналогичное явление наблюдалось при предъявлении проб, адресуемых к разным сенсорным входам (зрительным и слуховым) и с речевым ответом или без речевого ответа (рис. 33).
Из приведенных данных можно с учетом динамичности нейронных коррелятов импульсной активности сделать следующие осторожные заключения. В нейронных популяциях, расположенных в области бледного шара, обнаружены реакции как активирующего, так и тормозного типа. Эти реакции были сходными и различными при разных психологических пробах. При этом селективность реакций тормозного типа, если они наблюдаются в отведениях из бледного шара и передних отделов лобной коры, невелика. В нейронных популяциях, расположенных в области вентролатерального ядра зрительного бугра, в области теменной коры, в отдельных
популяциях бледного шара, обнаруживаются также реакции активирующего типа, однако они более или менее высокоселективны, различны при разных пробах.
Сравнение между собой реакций тормозного типа (см. выше) показывает, что урежение частоты импульсной активности, независимо от вида мыслительной деятельности, наблюдалось раньше в подкорке (бледный шар), чем в коре (передние отделы лобной коры).

Рис. 33. Реакции нейронов мозга человека в ответ на предъявление зрительных и слуховых стимулов.
Профили реакции всех обследованных нейронных популяций (слева) и ПСГ, вычисленные для отдельных нейронных популяций подкорковых образований мозга в ответ на зрительное (верхний ряд), слуховое (средний ряд) предъявления цифр в отсутствие вербальной реакции и в случае, когда человек называет цифру вслух (нижний ряд). По оси ординат на профилях реакций - число нейронных популяций, в которых были обнаружены значимые (Р < 0.05) отличия частоты разряда в бине от средней фоновой частоты, на ПСГ - средняя частота разряда в бине; по оси абсцисс - время в бинах (бин - 100 мс). На гистограммах: S —предъявление информационного стимула (цифры); S - предъявление триггерного стимула, служащего сигналом для вербальной реакции испытуемого; R - название стимула. Остальные обозначения - как на рис. 32
Эти данные, однако, должны расцениваться пока как вероятность появления такого рода нейродинамики. Только длительные повторные исследования позволят уточнить, насколько именно вероятна та или другая из описанных ситуаций. Ивто же время уже можно говорить о более или менее селективной реакции на психологические пробы в различных нейронных популяциях.

Рис. 34. ПСГ импульсной активности нейронной популяции в хвостатом ядре левого полушария при выполнении серии тестов, связанных со сравнением слов, квазислов, рисунков известных и неизвестных объектов.
Схематическое изображение структуры пробы дано в середине нижней части рисунка. В любой пробе предъявляются два стимула, каждый из которых может быть рисунком известного (РИС) или неизвестного (кв РИС) объекта, словом (СЛ) или квазисловом (кв СЛ). В зависимости от даваемой перед началом теста инструкции испытуемый в каждой пробе отвечает «Да» или «Нет» с учетом особенностей предъявленных стимулов
Большую или меньшую селективность реакций нейронных популяций удалось наблюдать и при предъявлении совокупностей тестов с опознанием слов, сравнением слов и квазислов, слов с соответствующими и не соответствующими им изображениями, сравнением изображений, имеющих и не имеющих смыслового значения, и т. п. Иллюстрацией сказанного может служить рис. 34 (Гоголицын и др., 19876).
Особую осторожность (опять с учетом динамичности нейронных коррелятов мышления) следует проявлять в этих исследованиях при оценке негативных эффектов
- когда нет значимых изменений импульсной активности при психологических пробах. При наличии перестроек на основе одного-двух тестов правильнее говорить, что данная зона, являясь звеном системы, может более или менее постоянно участвовать в обеспечении данного вида деятельности. При отсутствии перестроек для того, чтобы сказать, что данная зона мозга не может участвовать в обеспечении реализуемой деятельности, безусловно, необходимы повторные исследования (особенно это относится к случаям с высокой селективностью реакций). С учетом этих положений, может быть, следует осторожнее относиться к данным, полученным
в результате электрической стимуляции открытого мозга (Ojemann, 1983), на основе которых описывается высокая функциональная селективность в обеспечении мыслительных и преимущественно вербально-мнестических процессов.
Исследования импульсной активности нейронных популяций подтвердили многие механизмы мозга, показанные или предсказанные нами ранее. Эти работы безусловно явились и существенно новой ступенью в изучении различных мозговых механизмов. Это, в частности, относится к динамичности мозговых коррелятов мыслительной деятельности, которая была, во-первых, гораздо полнее изучена, доказана и, во- вторых, рассмотрена более полно в плане внутренних механизмов, определяющих динамичность нейрофизиологических коррелятов мыслительных процессов.
Ранее нами (Бехтерева и др. 1970) было показано, как могут изменяться свойства зоны мозга в зависимости от состояния различных видов биохимической медиации. Позднее В. А. Илюхиной (1977) была обнаружена зависимость свойств мозговых зон от уровня так называемого постоянного потенциала, наиболее стабильной составляющей СМФП - омега-потенциала (по: Илюхина, 19826). При одновременном отведении и исследовании импульсной активности нейронных популяций менее стабильных СМФП была обнаружена зависимость функциональной характеристики зоны мозга и от этого показателя.
В исследованиях С. В. Медведева и сотрудников (Медведев, Белов, 1985, 1986; Медведев и др., 1987) регистрировалась динамика импульсной активности нейронов (ИАН) при одновременном отведении с той же точки мозга, с того же электрода (что определялось возможностями полиэлектронейрографа, - Данько, Каминский, 1982) СМФП в декасекундном диапазоне (тау-волн, по В. А. Илюхиной). Больному предъявлялись психологические пробы на обобщение зрительного стимула.
В зонах мозга, расположенных в области различных ядер таламуса, стриопаллидарной системы была обнаружена зависимость динамики ИАН по ходу реализации психологического теста от фазовых характеристик тау-волн в той же зоне (рис. 35). Особенно интересной была в этом случае тонкая модуляция функциональных свойств мозговых зон.
Как представлено в работах В. А. Илюхиной (1977, 1986), при изменениях наиболее стабильной составляющей СМФП наблюдались перестройки функциональных свойств зон мозга, которые можно было характеризовать типом «да-нет», то есть свойство либо исчезало, либо появлялось. При изменениях фазы тауволн могло наблюдаться уменьшение или увеличение изменений ИАН в различные фазы психологического теста. Так, например, в зоне, расположенной в области заднего вентроорального ядра таламуса, при реализации психологического теста на восходящей и частично нисходящей фазе тау-волны существенные изменения ИАН обнаруживались в фазе принятия решения. В той же зоне при реализации того же теста на нисходящей-восходящей (рис. 35) фазе тау-волны изменения частоты ИАН в
фазу принятия решения отсутствовали.
При предъявлении в тесте двух абсолютно одинаковых сигналов, из которых только первый нес информационную нагрузку, а второй был в контексте теста незначим, на восходящей- нисходящей фазе тау-волны обнаруживалась реакция только на первый из двух сигналов. А при предъявлении их на нисходящей- восходящей фазе селективность реакции зоны мозга исчезала - зона реагировала одинаково на оба сигнала.

Рис. 35. Зависимость
динамики импульсной активности нейронов от фазовых характеристик тау-волн тех же зон мозга при мыслительной деятельности.
А - результаты усреднения СМФП в декасекундном диапазоне по положительным (б, г, е) и отрицательным (а, в, д) группам знакового ключа; Б - ПСГ текущей частоты импульсной активности нейрона: а, б, д, е - смысловые группы; в, г - «пустышки»; а- г - регистрация в области заднего вентроорального ядра таламуса (V. о. р); д, е - регистрация в области заднего дорсоорального ядра таламуса (D. о, р). Вертикальными штриховыми линиями обозначены моменты предъявления I (S), II (S) стимулов и сигнала, разрешающего речевой ответ больному (SR)