I. Спинной мозг.
Спинной мозг представляет собой низший и филогенетически наиболее древний отдел ЦНС. Спинной мозг характеризуется выраженным сегментарным строением, отражающим сегментарное строение тела позвоночных. Спинной мозг имеет многочисленные взаимные связи с головным мозгом. Оба этих отдела ЦНС в норме функционируют как единое целое. У млекопитающих, в частности у человека, сегментарная деятельность спинного мозга постоянно испытывает на себе влияние эфферентных нервных импульсов, исходящих из различных структур головного мозга. Это влияние в зависимости от многих обстоятельств может быть активизирующим, облегчающим или тормозящим.
Серое вещество спинного мозга составляют главным образом тела нервных и глиальных клеток. Разное количество их на разных уровнях спинного мозга обусловливает вариабельность объема и конфигурации серого вещества. В шейном отделе спинного мозга передние рога широкие, в грудном отделе серое вещество на поперечном срезе становится похожим на букву «Н», в пояснично-крестцовом отделе особенно значительны размеры как передних, так и задних рогов. Серое вещество спинного мозга фрагментируется на сегменты. Сегментом является фрагмент спинного мозга, анатомически и функционально связанный с одной парой спинномозговых нервов. Передние, задние и боковые рога можно рассматривать как фрагменты вертикально расположенных столбов - переднего, заднего и бокового, отделенных друг от друга состоящими из белого вещества канатиками спинного мозга.
В осуществлении рефлекторной деятельности спинного мозга важную роль играет следующее обстоятельство: практически все аксоны клеток спинномозговых узлов, входящие в спинной мозг в составе задних корешков, имеют ответвления - коллатерали. Коллатерали чувствительных волокон контактируют непосредственно с периферическими мотонейронами, расположенными в передних рогах, или с вставочными нейронами, аксоны которых также достигают тех же двигательных клеток. Коллатерали аксонов, отходящих от клеток межпозвонковых узлов, не только доходят до соответствующих периферичес- ких мотонейронов, расположенных в передних рогах ближайших сегментов спинного мозга, но и проникают в соседние его сегменты, формируя при этом так называемые спинно-спинальные межсегментарные связи,обеспечивающие иррадиацию возбуждения, пришедшего в спинной мозг после раздражения расположенных на периферии рецепторов глубокой и поверхностной чувствительности. Этим объясняется распространенная рефлекторная двигательная реакция в ответ на локальное раздражение.
Нервные клетки, составляющие серое вещество спинного мозга, по своей функции могут быть разделены на следующие группы:
| 1. Чувствительные клетки являются телами вторых нейронов чувствительных путей. Большая часть аксонов вторых нейронов чувствительных путей в составе белой спайки переходит на противоположную сторону, где участвует в формировании боковых канатиков спинного мозга, образуя в них восходящие спиноталамические пути и передний спиномозжечковый тракт Говерса. Аксоны вторых нейронов, не перешедшие на противоположную сторону, направляются в гомолатеральный боковой канатик и формируют в нем задний спиномозжечковый путь Флексига. |
2. Ассоциативные (вставочные) клетки, относящиеся к собственному аппарату спинного мозга, участвуют в формировании его сегментов. Их аксоны заканчиваются в сером веществе тех же или близко расположенных спинальных сегментов.
3. Вегетативные клетки расположены в боковых рогах спинного мозга на уровне CVIII - LII сегментов (симпатические клетки) и в сегментах SIII-SV (парасимпатические клетки). Аксоны их покидают спинной мозг в составе передних корешков.
4. Двигательные клетки (периферические мотонейроны) составляют передние рога спинного мозга. К ним сходится большое количество нервных импульсов, идущих из различных отделов головного мозга по многочисленным нисходящим пирамидным и экстрапирамидным путям. Кроме того, нервные импульсы к ним приходят по коллатералям аксонов псевдоуниполярных клеток, тела которых находятся в спинномозговых узлах, а также по коллатералям аксонов чувствительных клеток задних рогов и ассоциативных нейронов того же или других сегментов спинного мозга, несущих информацию главным образом от рецепторов глубокой чувствительности, и по аксонам, расположенным в передних рогах спинного мозга, клеток Реншоу, которые посылают импульсы, снижающие уровень возбуждения альфа-мотонейронов и, следовательно, уменьшающие напряжение поперечнополосатых мышц.
| Клетки передних рогов спинного мозга служат местом интеграции возбуждающих и тормозных импульсов, поступающих от различных источников. Среди периферических мотонейронов, расположенных в передних рогах спинного мозга, выделяются клетки двух видов: а) альфа-мотонейроны - крупные двигательные клетки, аксоны которых имеют толстую миелиновую оболочку (волокна А-альфа) и заканчиваются в мышце концевыми пластинками; они обеспечивают степень напряжения экстрафузальных мышечных во- локон, составляющих основную массу поперечнополосатых мышц; б) гамма-мотонейроны - мелкие двигательные клетки, аксоны которых имеют тонкую миелиновую оболочку (волокна А-гамма) и, следовательно, меньшую скорость проведения нервных импульсов. Гамма-мотонейроны составляют приблизительно 30% от всех клеток передних рогов спинного мозга; аксоны их направляются к интрафузальным мышечным волокнам, входящим в состав проприорецепторов - мышечных веретен. |
Таким образом, с участием гамма-клеток и их волокон создается гамма-петля, обеспечивающая поддержание тонуса мышцы и фиксированное положение определенной части тела или сокращение соответствующих мышц. Кроме того, гамма-петля обеспечивает трансформацию рефлекторной дуги в рефлекторное кольцо и принимает участие в формировании, в частности, сухожильных, или миотатических, рефлексов.
Моторные нейроны в передних рогах спинного мозга формируют группы, каждая из которых иннервирует мышцы, объединенные общностью функций. Передневнутренние группы клеток передних рогов, обеспечивающие функцию мышц, влияющих на положение позвоночного столба, и передненаружные группы периферических мотонейронов, от которых зависит функция остальных мышц шеи и туловища. В сегментах спинного мозга, обеспечивающих иннервацию конечностей, имеются дополнительные группы клеток, располагающиеся преимущественно позади и снаружи этих клеточных объединений. Эти дополнительные группы клеток являются основной причиной шейного (на уровне СV-ThII сегментов) и поясничного (на уровне LII-SII сегментов) утолщений спинного мозга. Они обеспечивают главным образом иннервацию мышц верхних и нижних конечностей.
Белое вещество спинного мозга формируется расположенными по периферии спинного мозга канатиками, состоящими из восходящих и нисходящих проводящих путей. Нервные волокна, имеющиеся в спинном мозге, можно дифференцировать на эндогенные, являющиеся отростками собственных клеток спинного мозга, и экзогенные - состоящие из проникших в спинной мозг отростков нервных клеток, тела которых расположены в спинномозговых узлах или входят в состав структур головного мозга. Эндогенные волокна могут быть короткими и длинными. Чем короче волокна, тем ближе к серому веществу спинного мозга они расположены. Короткие эндогенные волокна формируют спиноспинальные связи между сегментами самого спинного мозга. Из длинных эндогенных волокон, являющихся аксонами вторых чувствительных нейронов, тела которых расположены в задних рогах сегментов спинного мозга, сформированы афферентные пути, проводящие импульсы болевой и температурной чувствительности, идущие к таламусу, и импульсы, направляющиеся к мозжечку (спиноталамические и спиномозжечковые пути).
Экзогенные волокна спинного мозга являются аксонами клеток, находящихся за его пределами (рисунок 14). Они могут быть афферентными и эфферентными. Афферентные экзогенные волокна составляют тонкий и клиновидный пучки, формирующие задние канатики. Среди эфферентных проводящих путей, состоящих из экзогенных волокон, следует отметить боковой и передний кортико-спинальные тракты. Из экзогенных волокон состоят также относящиеся к экстрапирамидной системе красноядерно-спинномозговой, преддверно-спинномозговой, оливо-спинномозговой, покрышечно-спинномозговой, вестибу-лоспинномозговой, ретикуло-спинномозговой проводящие пути.
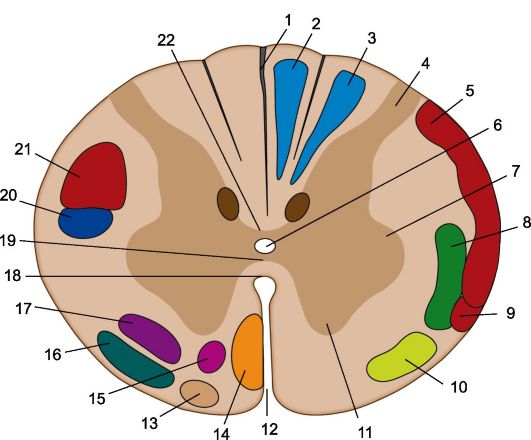
Рис. 14. Проводящие пути на поперечном срезе верхнегрудного отдела спинного мозга. 1 - задняя срединная перегородка; 2 - тонкий пучок; 3 - клиновидный пучок; 4 - задний рог; 5 - спиномозжечковый путь, 6 - центральный канал, 7 - боковой рог; 8 - латеральный спиноталамический путь; 9 - передний спиномозжечковый путь; 10 - передний спиноталамический путь; 11 - передний рог; 12 - передняя срединная щель; 13 - оливоспинномозговой путь; 14 - передний корково-спинномозговой (пирамидный) путь; 15 - передний ретикулярно-спинномозговой путь; 16 - преддверноспинномозговой путь; 17 - ретикулярно-спинномозговой путь; 18 - передняя белая спайка; 19 - серая спайка; 20 - красноядерно-спинномозговой путь; 21 - латеральный корково-спинномозговой (пирамидный) путь; 22 - задняя белая спайка.
СТВОЛ МОЗГА
В последние десятилетия к стволу мозга относят продолговатый мозг, варолиев мозг и средний мозг.
Ствол мозга имеет длину 8-9 см, ширину 3-4 см. Масса его невелика (30-32 г), однако его функциональное значение исключительно важно и многообразно, поскольку от расположенных в нем структур зависит жизнеспособность организма.
Если ствол мозга представить в горизонтальном положении, то на его сагиттальном срезе определяются 3 «этажа»: основание, покрышка, крыша.
Основание прилежит к скату затылочной кости. Его составляют нисходящие (эфферентные) проводящие пути (корково-спинальные, корковоядерные, корково-мостовые), а в мосту мозга - мосто-мозжечковые связи.
Покрышкой принято называть часть ствола, расположенную между его основанием и вместилищами спинномозговой жидкости (СМЖ) - четвертым желудочком, водопроводом мозга. Ее составляют двигательные и чувствительные ядра черепных нервов, красные ядра, черная субстанция, восходящие (афферентные) проводящие пути, в том числе спиноталамические пути, медиальная и латеральная петли и некоторые эфферентные экстрапирамидные пути, а также ретикулярная формация (РФ) ствола и их связи.
Крышей ствола мозга можно условно признать структуры, расположенные над проходящими через ствол вместилищами СМЖ. К ней относятся мозжечок, задний и передний мозговые паруса. Крышей среднего мозга называется пластинка четверохолмия.
Ствол мозга - продолжение верхнего отдела спинного мозга, сохраняющее элементы сегментарного строения. Как и в спинном мозге, серое вещество ствола расположено в глубине. Оно состоит из ретикулярной формации (РФ) и других клеточных структур, в него входят и ядра черепных нервов. Среди этих ядер различают двигательные, чувствительные и вегетативные. Условно их можно рассматривать как аналоги соответственно передних, задних и боковых рогов спинного мозга. Как в двигательных ядрах ствола, так и в передних рогах спинного мозга находятся двигательные периферические нейроны, в чувствительных ядрах - вторые нейроны путей различных видов чувствительности, а в вегетативных ядрах ствола, как и в боковых рогах спинного мозга, - вегетативные клетки.
II. Продолговатый мозг.
Продолговатый мозг - непосредственное продолжение спинного мозга. Условная граница между ними располагается на уровне большого затылочного отверстия; она проходит через первые спинномозговые корешки, или зону перекреста пирамидных путей. Продолговатый мозг имеет длину 2,5-3 см, по форме он похож на опрокинутый усеченный конус; иногда его называют луковицей (рисунок 15).Нижняя часть продолговатого мозга находится на уровне края большого затылочного отверстия, а верхняя, более широкая, граничит с мостом мозга. Условная граница между ними проходит на уровне середины ската затылочной кости.
 Рис. 15. Продолговатый мозг:
1–пирамиды; 1а–перекрест; 2–нижние оливы; 3–передний спинномозжечковый путь; 4– задний спинномозжечковый путь; 5–клиновидное ядро; 6–нежное ядро;7–; 8–ретикулярная формация; 9–вестибулоспинальный путь; 10–путь от бугров четверохолмия; 11–красноядерно-спинальный путь; 12–волокна межоливного слоя; 13–перекрест путей Голля и Бурдаха; IX–XII–ядра черепно-мозговых нервов
Рис. 15. Продолговатый мозг:
1–пирамиды; 1а–перекрест; 2–нижние оливы; 3–передний спинномозжечковый путь; 4– задний спинномозжечковый путь; 5–клиновидное ядро; 6–нежное ядро;7–; 8–ретикулярная формация; 9–вестибулоспинальный путь; 10–путь от бугров четверохолмия; 11–красноядерно-спинальный путь; 12–волокна межоливного слоя; 13–перекрест путей Голля и Бурдаха; IX–XII–ядра черепно-мозговых нервов
|
На вентральной поверхности продолговатого мозга в сагиттальной плоскости проходит глубокая продольная передняя срединная щель, являющаяся продолжением одноименной щели спинного мозга. По бокам от нее расположены возвышения - пирамиды, состоящие из корковоспинномозговых путей, включающих аксоны центральных мотонейронов. Позади и латеральнее пирамид с каждой стороны продолговатого мозга находится по нижней оливе. Из расположенной между пирамидой и оливой переднебоковой борозды выходят корешки подъязычного (XII) нерва. Позади олив находится задняя латеральная борозда,через которую из продолговатого мозга проходят корешки добавочного, блуждающего и языкоглоточного (XI, X и IX) нервов. В нижней части дорсальной поверхности продолговатого мозга между задней срединной бороздой и задними боковыми бороздами находятся два продольных валика, состоящих из пришедших сюда по задним канатикам спинного мозга волокнам нежного и клиновидного пучков.
Большую часть задней поверхности продолговатого мозга составляет нижний треугольник дна IV мозгового желудочка - ромбовидной ямки, которую снизу ограничивают нижние, а сверху - верхние ножки мозжечка.
Белое вещество продолговатого мозга состоит из проводящих путей, часть из которых проходит через него транзитом, часть прерывается в ядрах продолговатого мозга и входящей в его состав РФ или начинается от этих его структур. Корково-спинномозговые (пирамидные) пути проходят через основание продолговатого мозга, участвуя в формировании находящихся в его составе пирамид, и затем совершают неполный перекрест. Подвергшиеся перекресту волокна корково-спинномозгового пути сразу же попадают в состав латеральных канатиков спинного мозга; волокна же этого пути, не участвующие в формировании перекреста, включаются в состав переднего спинального канатика. Как перешедшие на противоположную сторону, так и оставшиеся на своей стороне волокна корково-спинномозгового пути, а также другие эфферентные связи, спускающиеся из различных структур головного мозга в спинной мозг, направляются к находящимся в передних рогах спинного мозга периферическим двигательным нейронам.
Продолговатый мозг посредством черепно–мозговых нервов (VIII – XII пары) связан с разнообразными рецепторами и эффекторами организма, а через восходящие и нисходящие проводящие пути – с другими отделами ЦНС.
Продолговатый мозг осуществляет рефлекторную и проводниковую функции. Рефлексы продолговатого мозга сложнее и разнообразнее рефлексов спинного. Функции продолговатого мозга имеют жизненно важное значение: в нем находятся дыхательный и сосудистый центры; продолговатый мозг оказывает вегетативное рефлекторное влияние на внутренние органы и сосуды (Х пара ч.м.н.). Кроме того, продолговатый мозг осуществляет ряд защитных рефлексов (мигание, чихание, кашель, рвотный акт и др.), ряд рефлексов, связанных с пищеварительной деятельностью (глотание, отделение пищеварительных соков и др.), и тонические рефлексы на скелетную мускулатуру.
Большой интерес вызывают рефлексы позы, осуществляемые с уровня продолговатого мозга.
В процессе эволюции у животного и человека выработалась нормальная поза, характеризующаяся положением головы теменем вверх. Поэтому у организма возникла и закрепилась группа тонических рефлексов, направленных на сохранение равновесия при угрозе его нарушения и на восстановление нормальной позы в тех случаях, когда она уже нарушена. Рефлексы позы возникают при изменении положения головы в пространстве и по отношению к туловищу. Так, при наклоне или при повороте головы в ту или другую сторону можно наблюдать рефлекторное увеличение тонуса разгибателей с соответствующей стороны и снижение его на противоположной. При опускании головы животного происходит снижение разгибательного тонуса передних конечностей при некотором повышении тонуса задних. При подъеме головы кверху происходит повышение тонуса мышц-разгибателей передних конечностей и снижение – на задних.
Рефлексы позы осуществляются с рецепторов вестибулярного аппарата, с проприорецепторов мышц и сухожилий шеи, а также с экстерорецепторов кожи шеи.
Отчетливое проявление позных рефлексов можно наблюдать на больных людях при нарушении у них деятельности вышележащих отделов мозга. У здорового человека при изменении положения головы методом рентгенокимографии было обнаружено повышение тонуса диафрагмы с той стороны, куда была повернута голова.
III. Средний мозг.
Средний мозгможно рассматривать как продолжение моста и верхнего переднего паруса. Он имеет длину в 1,5 см, состоит из ножек мозгаи крыши, или пластинки четверохолмия. Условная граница между крышей и подлежащей покрышкой среднего мозга проходит на уровне водопровода мозга (сильвиев водопровод), который является полостью среднего мозга и соединяет между собой III и IV желудочки мозга.
Ножки мозга хорошо видны на вентральной стороне ствола. Они представляют собой два толстых тяжа, которые выходят из вещества моста и, постепенно расходясь в стороны, вступают в большие полушария головного мозга. В том месте, где ножки мозга отходят друг от друга, между ними находится межножковая ямка, закрытая так называемым задним продырявленным веществом.
Основание среднего мозга образуют вентральные отделы ножек мозга. В отличие от основания моста здесь нет поперечно расположенных нервных волокон и клеточных скоплений. Основание среднего мозга составляют только продольные эфферентные пути, идущие из больших полушарий через средний мозг к расположенным ниже отделам ствола и к спинному мозгу. Лишь небольшая их часть, входящая в состав корково-ядерного проводящего пути, заканчивается в покрышке среднего мозга, в расположенных здесь ядрах III и IV черепных нервов (рисунок 16).

Рис. 16. Средний мозг
I – крыша среднего мозга; II – покрышка среднего мозга; III – основание среднего мозга; 1 – верхние холмики; 2 – ядро верхнего холмика; 3 – серое вещество среднего мозга; 4 – ядро среднего мозга (V); 5 - водопровод; 6 – глазодвигательное ядро; 7 – ядро Якубовича; 8 – ядро Перлея; 9 – кортико-мостовой путь; 10,11 – кортикоспинальные пути; 12 – кортико-мостовой путь; 13 – черная субстанция; 14 – дорсальный перекрест Мейнерта; 15 – вентральный перекрест Фореля; 16 – красное ядро; 17 – ретикулярная формация; 18 – медиальная петля; 19 – боковая петля; 20 – ядро нижнего холмика
Волокна, составляющие основание среднего мозга, расположены в определенном порядке. Среднюю часть (3/5) основания каждой ножки мозга составляют пирамидные и корково-ядерные проводящие пути; медиальнее от них расположены волокна лобно-мостового пути Арнольда; латеральнее - волокна, идущие к ядрам моста из теменной, височной и затылочной долей больших полушарий мозга, - путь Тюрка.
Над этими пучками эфферентных проводящих путей расположены структуры покрышки среднего мозга, содержащие ядра IV и III черепных нервов, парные образования, относящиеся к экстрапирамидной системе (черная субстанция и красные ядра), а также структуры ретикулярной формации, фрагменты медиальных продольных пучков, а также многочисленные проводящие пути различной направленности.
Между покрышкой и крышей среднего мозга располагается узкая полость, имеющая сагиттальную направленность и обеспечивающая сообщение между III и IV мозговыми желудочками, именуемая водопроводом мозга.
Средний мозг имеет «собственную» крышу - пластинку четверохолмия, в состав которой входят два нижних и два верхних холмика. Задние холмики относятся к слуховой ориентировочной системе, передние - к зрительной.
Черная субстанция участвует в сложной координации движений. Ее нейроны, содержащие дофамин, своими аксонами связаны с передним мозгом (участие в регуляции эмоционального поведения), и ядрами полосатого тела (контроль сложных двигательных актов). Повреждения черной субстанции приводят к нарушению тонких движений пальцев рук, развитию мышечной ригидности и тремору (болезнь Паркинсона).
В составе среднего мозга также имеются важные эффекторные образования – красные ядра. Они принимают значительное участие в осуществлении мышечного тонуса и нормальной позы тела. При перерезке ствола мозга ниже уровня красного ядра у животных наблюдается резкое повышение тонуса мышц разгибателей. Конечности при этом сильно вытянуты, голова запрокинута назад, хвост приподнят. Это состояние называется децеребрационная ригидность. Развитие децеребрационной ригидности связано с выключением импульсации, поступающей в спинной мозг по кортико–, ретикуло– и руброспинальному трактам, которые активируют мотонейроны мышц–сгибателей. При этом начинает преобладать активность вестибулоспинальной системы, повышающей тонус мотонейронов мышц–разгибателей. Децеребрационная ригидность наблюдается и у человека при травмах среднего мозга.
Средний мозг осуществляет также ряд установочных тонических рефлексов – выпрямительных и статокинетических