Явление, когда признак развивается при взаимодополняющем дей-ствии доминантных неаллельных генов. По типу комплементарности взаимо-действуют гены, контролирующие разные этапы одного и того же метаболи-ческого пути. Впервые такой тип взаимодействия генов был описан Бетсо-ном и Пеннетом при анализе наследования окраски венчика у душистого горошка. При скрещивании растений душистого горошка с белыми цветами в первом поколении все растения имели пурпурные цветы (как у дикого вида душистого горошка) формы, а во втором поколении наблюдалось расщепле-нии в соотношении 9 пурпурные: 7 белые.
Для объяснения полученного результаты было сделано предположе-ние, что синтез пурпурного пигмента контролируют два неаллельных доми-нантных гена, продукты которых задействованы на разных этапах метаболического пути. Мутация любого из этой пары гена прерывает процесс синтеза пигмента. Взятые для скрещивания растения в своем генотипе имели мутации разных комплементарных генов.

Схема скрещивания:
Р: белые венчики белые венчики
♀ ААbb х ♂ ааВВ
G: Аb аВ
F1: АаВb – 100% пурпурные венчики
F2: 9/16 – пурпурные (А•В•): 7/16 белые (3/16 – А•bb, 3/16 – ааВ•; 1/16 – ааbb).
Вторым примером взаимодополняющего действия может служить наследование окраски шерсти у мышей (рис. 41), когда при скрещивании чи-стопородных линий черных и белых мышей, все гибриды первого поколения (F1) имеют серую окраску шерсти (комплементарная), а в F2 наблюдается расщепление по фенотипу 9 серые: 3 черные: 4 белые.
При этом первая пара аллельных генов отвечает за синтез пигмента (С – синтез пигмента; с – отсутствие пигмента (альбинизм)), вторая пара генов – за распределение пигмента по длине волоса (А – зональное распределение пигмента; а – не определяет зонального распределения пигмента).
| Формула дигибридного расщепления по фенотипу: |
| 9: 3: 3: 1 |
| А•C• ааC• А•cc ааcc |
| серая черная белая белая |
| 9: 3: 4 |
Третьим примером взаимодополняющего действия является наследо-вание формы плода у тыквы (рис.42). В результате комплементарного взаи-модействия генов в F2 наблюдается соотношение 9: 6: 1.

Комплементарность (комбинативное взаимодействие)
Один из первых примеров взаимодействия неаллельных генов был об-наружен в начале XX в. при анализе наследования формы гребня у кур. Опи-сано четыре разновидности форм гребней, при этом разные породы имеют характерную морфологию гребня (рис. 43):

В результате скрещиваний кур, имеющих розовидный и гороховидный гребни, в F1 возникает новая форма гребня – ореховидный (из-за взаимодей-ствия генов A и B).
Р: Розовидный Гороховидный
♀ AAbb х ♂ aaBB
G: Ab aB
F1: AaBb – 100% Ореховидный
Скрещивание гибридов F1 дает следующие результаты в F2:
| AB | Ab | aB | Ab | |
| AB | Орех. AABB | Орех. AABb | Орех. AaBB | Орех. AaBb |
| Ab | Орех. ААВb | Розов. ААbb | Орех. АаВb | Розов. Ааbb |
| аB | Орех. АаВВ | Орех. АаВb | Горох. ааВВ | Горох. ааВb |
| аb | Орех. АаВb | Розов. Ааbb | Горох. ааВb | Листов. ааbb |
ЭПИСТАЗ (подавление) – ген одной аллельной пары подавляет дей-ствие генов другой аллельной пары. Ген, подавляющий действие другого гена, называется эпистатическим геном, ингибитором или супрессором. Подавляемый ген носит название гипостатического.
| ЭПИСТАЗ (расщепление по фенотипу) | |
| F2: | 13: 3 |
| 12: 3: 1 | |
| 9: 3: 4 |
Различают доминантный и рецессивный эпистаз.
При доминантном эпистазе доминантный аллель одного гена (ген-супрессор) подавляет действие доминантной или рецессивной аллели другого гена (гипостатический ген). Ген А (супрессор) подавляет ген В (гипостатический) или ген А (эпистатический) подавляет ген b (гипостатический). При доминантном эпистазе в дигибридном скрещивании происходит расщепление в соотношении 12:3:1 или 13:3.
Например, при скрещивании 2 белых кур леггорн ААСС и виандотт аасс во втором поколении произойдёт расщепление 13 белых: 3 окрашен-ных. Ген С подавляет ген А. В отсутствии гена С проявляется ген А, и куры окрашены (рис. 46).
У лошадей ген В обусловливает синтез черного пигмента (вороная масть), а ген b – рыжую масть. Ген С – эпистатический ген (серая масть – результат эпистаза), а его рецессивный аллель с не подавляет проявлений гена B и b. Лошади серой масти имеют генотип – 9 •ѕ и 3 bbС•; вороные – 3В•сс, рыжие – 1bbсс. Расщепление 12: 3: 1 (рис. 47).
В случае рецессивного эпистаза рецессивный эпистатический аллель в гомозиготном состоянии препятствует проявлению доминантной или ре-цессивной аллели другого гена (аа подавляет ген В или аа подавляет ген b). При рецессивном эпистазе в дигибридном скрещивании происходит расщеп-ление в соотношении 9: 3: 4.
Ярким примером рецессивного эпистаза у человека является "Бомбей-ский феномен". Так, у человека группы крови АВО контролируются тремя аллелями одного гена IA – вторая группа (синтез антигена А), IB – третья группа крови (синтез антигена В), i – первая группа кровит (синтез антигенов не происходит). В популяции встречается редкий мутантный аллель h неза-висимого гена, который в гомозиготном состоянии подавляет действие алле-лей А и В, что приводит к фенотипическому проявлению первой группы кро-ви.

В Индии была описана семья, в которой родители имели вторую и первую группу крови. Оба родителя были гомозиготными по группам крови, и рождение ребенка с четвертой группой крови в их семье вызвало недоуме-ние (рис. 49).
Таким образом, "Бомбейский феномен" определяется тем, что в гено-типе детей есть ген подавляющий действие генов IА и IВ.

Группа крови АВ0 хорошо исследована биохимически. Известно, что антигены А и В представляют собой карбогидратные группы (углеводные остатки), связанные с жирными кислотами, молекулы которых выступают над поверхностью мембраны эритроцитов. Специфичность этих антигенов обусловлена концевым остатком карбогидратной цепи. Оба антигена проис-ходят из одного и того же предшественника, или Н-вещества, к молекулам которого затем добавляется один или два концевых углеводных остатка сахара.
В крайне редких случаях, например, у женщин из Бомбея (Индия) имеется дефектное Н-вещество, которое не связывается с ферментом, добав-ляющим концевые углеводные остатки. У носителей группы крови 0 это при-водит к фенотипу Бомбей. Оказалось, что этот дефект обусловлен рецессив-ной мутацией h в локусе, не гомологичном локусам А и В. Поэтому при наличии у человека аллелей IA и (или) IB вместе с генотипом hh антигены А и В в крови не обнаруживаются. Вот почему у одной из жительниц Бомбея с генотипом hh определили группу крови 0, хотя ее родители имели группу крови АВ, а сын был носителем аллеля IB (рис. 50).
ПОЛИМЕРИЯ – явление, когда несколько неаллельных доминантных генов контролируют развитие одного и того же признака. Такие гены назы-ваются полимерными и обозначаются одной и той же буквой, но с другим индексом (например: А1, А2, А3, А4 и т.д.). Различают кумулятивную и неку-мулятивную полимерию.

При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество до-минантных аллелей не влияет на степень выраженности признака. Впервые некумулятивная полимерия была описана в 1908 г. Нильсоном-Эле при ана-лизе наследования окраски эндосперма зерновок пшеницы (рис. 51). Доми-нантные неаллельные гены А1 и А2 обуславливают красную окраску зерен пшеницы, их рецессивные аллели а1 и а2 – отсутствие окраски. Действие ге-нов А1 и А2 однозначно.
При наличии двух полимерных генов расщепление во втором поколе-нии соответствует 15: 1, при наличии трёх полимерных генов – 63: 1 и т.д.).
При кумулятивной полимерии действие полимерных генов суммиру-ется, чем их больше, тем ярче выражен признак. Расщепление во втором по-колении будет зависеть от количества полимерных генов (суммарный или аддитивный эффект генов), присутствующих в генотипе.
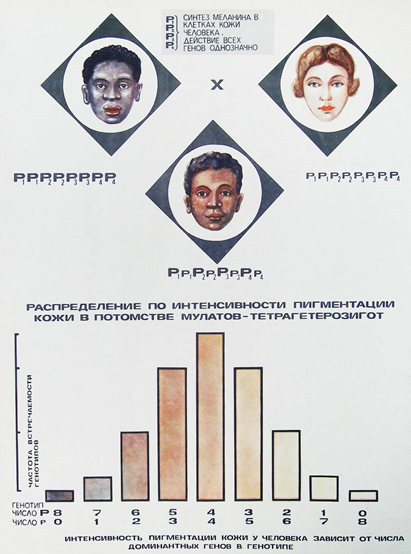
По типу кумулятивной полимерии наследуются многие количествен-ные признаки (у человека: пигментация кожи, рост, вес, уровень интеллекта, скорость протекания биохимических реакций, артериальное давление, со-держание сахара в крови; у животных: жирность молока; у растений: яро-вость, озимость, содержание витаминов, длина колоса у злаков, интенсив-ность окраски частей растения и др.).
Цвет кожи человека определяется взаимодействием нескольких пар генов по типу кумулятивной полимерии, т.е. цвет кожи тем темнее, чем больше доминантных генов в генотипе (рис. 52).

ЭФФЕКТ ПОЛОЖЕНИЯ ГЕНОВ – влияние расположения генов в хромосоме на проявление их активности. Явление открыто американским генетиком А. Стёртевантом в 1925 году. Наблюдается при структурных перестройках хромосом (транслокациях), в результате которых гены актив-ных зон хромосом (эухроматина) могут переноситься в неактивные зоны (ге-терохроматин) и инактивироваться и наоборот. При перестройке, возвраща-ющей эухроматиновый ген из гетерохроматина в любую точку эухроматина, функционирование данного гена восстанавливается.
Эффект положения гена можно проследить при наследовании резус фактора (Rh-фактор).
В простейшем варианте резус-положительность доминирует над резус-отрицательностью и практически наследование Rh-фактора иммитирует моногогенное наследование. Однако ряд исследований показывает, что система Rh определяется тремя антигенными факторами, которые детермируются тремя тесно сцепленными генами С, Д, Е, локализованными в коротком плече хромосомы 1. Внутри аллельных пар действует полное доминирование, а при взаимодействии между ними «эффект положения». Все это обусловливает разнообразие вариантов резус-антигенов и соответственно резус-несовместимости.
Основная роль в Rh-системе принадлежит антигену D. При его нали-чии на поверхности эритроцитов кровь является резус-положительной. Резус-отрицательный фенотип формируется при отсутствии антигена D.
Однако антигены C и E оказывают влияние на резус-несовместимость.
Суть «эффекта положения» в следующем: в рассмотренном примере (рис. 53) оба человека имеют одинаковые фенотипы и генотип. Они гетерози-готы по трем парам генов (CcDdEe) и оба резус-положительные, но в анти-генном плане их кровь несовместима. У человека, у которого в одной хромо-соме порядок расположения генов Cde, а в другой соответственно cDE, будут синтезироваться все три вида антигенов C, D, E и его кровь будет несовме-стима с кровью второго человека.
У второго человека порядок расположения генов внутри хромосомы cDe, а в гомологичной – CdE. Там, где в одной хромосоме расположены до-минантные гены С и Е, ген Е выступает в роли супрессора по отношению к гену С. Следовательно, фенотипическое проявление гена С в признак подав-лено, антиген С не синтезируется.
РЕЗУС-ФАКТОР И РЕЗУС-КОНФЛИКТ
Резус-фактор крови – антиген, представляющий собой особый вид про-теина, присутствующего на оболочке красных кровяных телец человека и макаки-резус. Открыт белок был в 1940 году А. Винером и К. Ландштейне-ром.
Этот белок есть у 85% жителей планеты европеоидной расы. Таких людей называют резус-положительными. Если же белка на оболочке эритро-цитов нет – они называются резус-отрицательными. Среди людей монголо-идной расы резус-отрицательных людей еще меньше – всего 1%, а среди жи-телей планеты негроидной расы только 7%.
Резус крови – это достаточно непростая конструкция, в которую вхо-дит не меньше сорока различных антигенов. Каждый из антигенов имеет свое «имя», которое прописывается с помощью букв и цифр. Наиболее распро-страненными антигенами являются Д, С, Е. Именно эти антигены наиболее активны.
Rh-пол. Rh-пол.
♀ Rr x ♂ Rr
R, r R, r
Rh-пол. Rh-отр.
RR; Rr; Rr; rr
75% 25%
Резус крови передаётся по наследству как доминантный признак (R – ген наличия резус фактора, а r – отсутствие его) и не меняется в течение всей жизни.
Если у родителей присутствует резус (Rr, Rr), то у ребенка резус мо-жет присутствовать (RR, Rr), а может и отсутствовать (rr).
Если один из родителей резус-положителен (RR, Rr), а второй отрица-телен (rr), – малыш может унаследовать как положительный резус (Rr), так и отрицательный (rr).
В том случае, если и у мамы и у папы резус отрицателен, ребенок обя-зательно также родится резус-отрицательным.
Обычно отрицательный резус-фактор никаких неприятностей его хозя-ину не приносит. Особого внимания и ухода требуют лишь резус-отрицательные беременные женщины.
Если у будущей мамы резус отрицательный, а у будущего папы поло-жительный, возникает опасность резус-конфликта. Но начаться он может лишь в том случае, если ребенок унаследует резус отца. Тогда его кровь бу-дет плохо совместима с кровью матери (рис. 54).
Во время беременности мать и плод едины, и, несмотря на то, что кровь их не смешивается, многие продукты обмена веществ да и отдельные клетки от плода поступают к матери, и наоборот. Этот обмен идет через структуры плаценты, через ту ее часть, которая называется плацентарным барьером.
Rh-кoнфликт возникает не ранее 7-8-й недели беременности, когда начинается формирование кроветворения у зародыша. Резус-положительные эритроциты плода преодолевают плацентарный барьер и, попадая в кровь матери, а ее организм, воспринимаются как нечто чужеродное. Организм матери начинает вырабатывать защитные антитела. Защищая мать, эти антитела несут серьезную угрозу ее ребенку. При тяжелой форме резус-конфликта возможна внутриутробная гибель плода и выкидыш на любом сроке беременности.
Антитела матери проникают через плаценту и разрушают эритроциты ребенка. В крови появляется большое количество вещества, называемого билирубином. Билирубин окрашивает кожу малыша в желтый цвет. Поскольку эритроциты плода непрерывно уничтожаются, его печень и селезенка стараются ускорить выработку новых эритроцитов, увеличиваясь при этом в размерах. В конце концов, и они не справляются с восполнением убыли эритроцитов. Возникает состояние анемии (низкое содержание в крови эритроцитов, гемоглобина). Резус-конфликт может быть причиной поражения головного мозга ребенка, нарушения функции слуха и речи. В самых тяжелых случаях резус-конфликт проявляется врожденной водянкой (отеком) плода, которая может привести к его гибели.
В тяжелых случаях новорожденному может помочь заменное перели-вание крови. Ему вводят одногруппную резус-отрицательную кровь и прово-дят реанимационные мероприятия.
Рис. 55. Гемолиз эритроцитов при резус-конфликте.
Учёт резус фактора, как и группы крови, является обязательным при переливании крови. Если в тело резус-отрицательного пациента влить кровь с присутствующим резусом, в организме начнут вырабатываться антитела (рис. 55), под действием которых наступает гемолиз эритроцитов донора (эритроциты, обладающие резусом, склеиваются в длинные цилиндры).
Для профилактики подобных осложнений необходимо переливать ре-зус-отрицательному реципиенту только резус-отрицательную кровь и перед переливанием производить пробу на совместимость не только по группам крови, но и по резус-фактору.
Изменчивость – это универсальное свойство живых организмов при-обретать новые признаки в процессе индивидуального развития организмов (онтогенеза).
Ненаследственная (фенотипическая, модификационная) изменчивость – это фенотипические различия у генетически одинаковых особей, спо-собность организмов изменять свой фенотип под влиянием различных фак-торов.
Причиной модификационной изменчивости являются изменения внешней среды обитания организма или его внутренней среды, не затрагива-ющие гены организма, наследственный материал и не вызывающие измене-ний генотипа особи.
Модификационная изменчивость является определенной, то есть все-гда соответствует факторам, которые ее вызывают.
Генотипическая изменчивость – при генотипической изменчивости происходит изменение наследственного материала, и обычно эти изменения наследуются (наследственная). Это основа разнообразия живых организмов.
Ч. Дарвин определял наследственную изменчивость как индивидуаль-ную и неопределенную.
Неопределенная изменчивость – это "бесконечно разнообразные не-значительные особенности, которыми отличаются особи того же вида и которые невозможно объяснить унаследованием их от одного из родителей или от более отдаленных предков". Таковые, по Дарвину, есть "резковыраженные отличия, проявляющиеся у молоди одного помета и у семян из одной и той же коробочки".
Индивидуальные различия – это многочисленные незначительные раз-личия, появляющиеся в потомстве одних и тех же родителей или у особей одного и того же вида, обитающих в одной и той же ограниченной местно-сти. Особи одного вида, благодаря индивидуальным отличиям, различаются по многим признакам.
Комбинативная изменчивость основывается на возникновении но-вых комбинаций генов родителей.