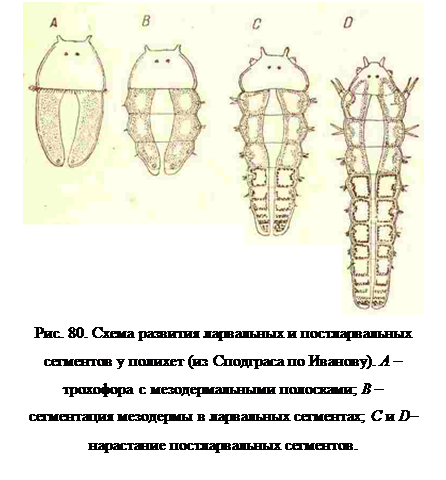
Метаморфоз трохофоры во взрослого червя у Polychaeta происходит путем разрастания посторального полушария ее в направлении продольной оси. Сначала разрастается и длину вся соматическая пластинка (производное 2 d и4 d). Этот рост закапчивается образованием червевидного тела, голова которого образована преоральным полушарном трохофоры, а само тело разделено на определенное в каждом семействе небольшое число сегментов (рис. 69, В, 70 и 80, А и В), в связи с образованием соответствующего имчисла пар параподий со щетинками (от 3 до 9 пар). Этот отдел у некоторых видов представляет собой тело сегментированной личинки или метатрохофоры (также нектохэты), и потому мы будем называть его сегменты ларвальными.
Постепенно или путем резкого перехода рост соматической пластинки или ларвального отдела сосредоточивается в ее заднем конце, непосредственно впереди анального отверстия, где возникает постоянная зона нарастания в длину, и образование все новых сегментов и пар нараподий доходит до очень большого числа (рис. 69, С и 80, D). Этот отдел тела, мы будем называть постларвальным.
Как в ларвальном, так и в постларвальном отделах одновременно с ростом идет процесс развития целомической мезодермы, первый зачаток которой имелся еще в трохофоре в виде пары первичных мезодермальных клеток, производных 4 d. Из них размножением клеток образуется пара сплошных мезодермальных полосок, тянущихся на всем протяжении ларвального отдела (рис. 80, А). Ларвальный отдел, достигнув в своем росте определенной длины, сразу сегментируется на все присущее данному семейству число ларвальных сегментов, и тогда на протяжении, всей мезодермальной полоски, путем расхождения ее клеток к эктодерме и к кишечнику и распределения их по сегментам тела, одновременно появляется ряд целомических полостей ларвальных сегментов (рис.80, В). На образование мезодермы ларвальных сегментов у одних Polychaeta расходуется весь материал первичных мезодермальных клеток и мезодермальных полосок, у других — только часть этого материала.
Образование постларвальных сегментов из концевой или телобластической зоны нарастания происходит последовательно, один сегмент за другим (рис. 80, С и D) так, что ближайшие к зоне роста сегменты представляют все последовательные переходные фазы своего формирования. Здесь в зоне роста возникает эктодерма, а по многих случаях вместе с нею и мезодерма этих сегментов. Мезодерма постларвальных сегментов возникает из клеток, оставшихся неизрасходованными на образование мезодермы ларвальныхсегментов. Если же весь этот материал оказался израсходованным, то эктодермальная зона роста дает также мезодерму.
 Образование отдельных мезодермальных сегментов или сомитов происходит в постларвальных сегментах совсем иначе, чем в ларвальных. Впереди концевого скопления интенсивно размножающихся клеток зоны роста отщепляется многослойная клеточная пластинка, справа и слева от кишечника, прикрепленная одним краем к этому последнему, а другим к стенке тела (рис. 80, С и D). В это время продолжается рост тела, вследствие чего зона роста отодвигается несколько назад, а от нее отщепляется вперед вторая такая же пластинка, в то время как первая несколько растягивается в поперечном к оси тела направлении. Таким же путем появляются следующие пластинки, или поперечные перегородки, отвечающие по своему положению будущим мезодермальным септам сегментов. Превращение такой утолщенной перегородки в сомит, т. е. замкнутыйцеломический мезодермальный сегмент, происходит путем сползания с нее клеток на стенку тела, где они образуют соматоплевру, и париетальный листок, и на кишечник, где они образуют спланхноплевру, или висцеральный листок. Так как при этом и париетальный и висцеральный слои достигают лежащей впереди септы, то получается замкнутая мешком мезодермальная выстилка, составляющая стенки сомита (рис. 80, С и D). Особому способу закладки мезодермы отвечают и анатомические особенности постларвальных сомитов; например, только в них имеются половые железы и обычные метанефридии.
Образование отдельных мезодермальных сегментов или сомитов происходит в постларвальных сегментах совсем иначе, чем в ларвальных. Впереди концевого скопления интенсивно размножающихся клеток зоны роста отщепляется многослойная клеточная пластинка, справа и слева от кишечника, прикрепленная одним краем к этому последнему, а другим к стенке тела (рис. 80, С и D). В это время продолжается рост тела, вследствие чего зона роста отодвигается несколько назад, а от нее отщепляется вперед вторая такая же пластинка, в то время как первая несколько растягивается в поперечном к оси тела направлении. Таким же путем появляются следующие пластинки, или поперечные перегородки, отвечающие по своему положению будущим мезодермальным септам сегментов. Превращение такой утолщенной перегородки в сомит, т. е. замкнутыйцеломический мезодермальный сегмент, происходит путем сползания с нее клеток на стенку тела, где они образуют соматоплевру, и париетальный листок, и на кишечник, где они образуют спланхноплевру, или висцеральный листок. Так как при этом и париетальный и висцеральный слои достигают лежащей впереди септы, то получается замкнутая мешком мезодермальная выстилка, составляющая стенки сомита (рис. 80, С и D). Особому способу закладки мезодермы отвечают и анатомические особенности постларвальных сомитов; например, только в них имеются половые железы и обычные метанефридии.
Проеоральное, или верхнее, полушарие трохофоры во время метаморфоза сначала заметно уменьшается и размерах; мерцательный же венец и теменной орган атрофируются. В это время начинается в предротовом отделе диференцировка надглоточного ганглия и органов чувств, после чего этот отдел с находящимися в нем зачатками растет, но гораздо слабее, чем остальное тело, так что у сформированного червя относительные размеры образующейся впереди рта предротовой лопасти становятся незначительными. Из органов чувств в головной или предротовой лопасти возникают глаза, обонятельные ямки, антенны и пальпы.
При наличии в яйце значительного количества желтка из яйца прямо выходит метатрохофора с полным числом вполне развитых ларвальных сегментов (рис. 81, Е, F). Дробление идет такжe спирально, но с некоторыми особенностями, вызнанными большой разницей в величине анимальных и вегетативных бластомер. После образования трех квартетов бластомеры основного квартета оказываются настолько набитыми желтком и бедными активной полярной протоплазмой, что, например,у Nereis о Capitella., четвертый квартет не образуется и главная масса полярной протоплазмы, оставшейся после перехода почти половины ее в 2 d попадает в 3 d, которая и становится материнской бластомерой для пары первичных мезодермальных клеток (рис. 81. В). Основной квартет послеобразования третьего квартета перестает делиться, служит лишь вместилищем желтка и вообще остается пассивным. Поэтому гаструляция происходит эниболией, и клетки трех квартетов располагаются по всей поверхности комплекса желточных  бластомер, где и происходит дальнейшее, формирование частей трохофоры. Ядра желточных бластомер могут при этом перемещаться и вместе с небольшими участками окружающей их протоплазмы функционируют как клетки, перерабатывающие желток, т. е. становятся и желточной энтодермой; энтодерма же кишечника возникает из мелких клеток (вероятно, отвечающих четвертому квартету или энтеробластам, отделяющимся при таком развитии от клеток 3 d. В таком случае расположение клеточного материала в всех частей тела трохофоры становится очень сходным с расположением частей зародыша при поверхностном дроблении членистоногих и части зародыша так же распластаны по поверхности желтка. Среди них видны и клетки трохобласты (рис. 81, С) и клетки теменного органа, по затем появляются зачатки параподиев всех трех ларвальных сегментов (у Nereis, рис. 81, D и Е), или девяти (у Capitеllidae), и к моменту выхода из яйца зародыша имеет уже все ларвальные папаодии и сегменты, а также мозг и головные, органы чувств.
бластомер, где и происходит дальнейшее, формирование частей трохофоры. Ядра желточных бластомер могут при этом перемещаться и вместе с небольшими участками окружающей их протоплазмы функционируют как клетки, перерабатывающие желток, т. е. становятся и желточной энтодермой; энтодерма же кишечника возникает из мелких клеток (вероятно, отвечающих четвертому квартету или энтеробластам, отделяющимся при таком развитии от клеток 3 d. В таком случае расположение клеточного материала в всех частей тела трохофоры становится очень сходным с расположением частей зародыша при поверхностном дроблении членистоногих и части зародыша так же распластаны по поверхности желтка. Среди них видны и клетки трохобласты (рис. 81, С) и клетки теменного органа, по затем появляются зачатки параподиев всех трех ларвальных сегментов (у Nereis, рис. 81, D и Е), или девяти (у Capitеllidae), и к моменту выхода из яйца зародыша имеет уже все ларвальные папаодии и сегменты, а также мозг и головные, органы чувств.
 Среди морских аннелид имеются, однако, формы, у которых после разделения тела трохофоры на ларвальные сегменты со слабо развитой в них мезодермой образуется только одни задний сегмент с хорошо оформленным целомом, в котором развиваются половые продукты и который, следовательно, отвечает всему многосегментному постларвальному отделу других аннелид. Таковы, например, Dinophilida (рис. 82). У паразитических Myzostomida тело также состоит из одних ларвальных сегментов. С другой стороны, имеются формы, у которых ларвальные сегменты их выражены, и на нижнем полюсе трохофоры прямо возникает зона нарастания постларвальных сегментов.Таков Poligodius(рис. 83) в близкие к нему Protodrilus н Saccocirrus.
Среди морских аннелид имеются, однако, формы, у которых после разделения тела трохофоры на ларвальные сегменты со слабо развитой в них мезодермой образуется только одни задний сегмент с хорошо оформленным целомом, в котором развиваются половые продукты и который, следовательно, отвечает всему многосегментному постларвальному отделу других аннелид. Таковы, например, Dinophilida (рис. 82). У паразитических Myzostomida тело также состоит из одних ларвальных сегментов. С другой стороны, имеются формы, у которых ларвальные сегменты их выражены, и на нижнем полюсе трохофоры прямо возникает зона нарастания постларвальных сегментов.Таков Poligodius(рис. 83) в близкие к нему Protodrilus н Saccocirrus.