У хитонов (Polyplarophora) личинка типичная трохофора. внутри которой, однако, уже хорошо развиты мезодермальные полоски и отсутствуют личиночные протонефридии. При метаморфозе на вентральной стороне возникает нога, в виде плоскойподошвы (рис. 103), а на спинной стороне появляются 7 светлых поперечных полос, представляющих собой зачатки пластинок раковины. Полоски становятся заметными вследствие чередования полосок высокого эпителия (рис. 104),выделяющего толстую кутикулу с более широкими полосками более плоского эпителия, выделяющего более тонкую кутикулу, над которой выделяется довольно толстый слой извести (рис. 105). Таким образом, получаются 7 поперечных пластинок раковины, отделенных друг от друга валиками высокого эпителия, а несколько позже сзади появляется еще, восьмая пластинка. Все пластинки вместе отвечают раковинкой железе, мантии и раковине других моллюсков. С появлением ноги и раковины личинка падает на дно и начинает вести образ жизни взрослого хитона. Появляется характерный для всех моллюсков зачаток терки, или radula, возникающий в виде глубокою впячения нейтральной стенки передней части кишечника.

 Вследствие расчленения раковины на метамерные пластинки, у хитонов метамерии группируются и мышцы тела и кровеносные сосуды; другими словами, мезодермальиые полоски их получают сегментировку, зависящую от метамерии эктодермальных образований, т. е. такую же. какая возникает в мезодермальных полосках ларвальных сегментов аннелид.
Вследствие расчленения раковины на метамерные пластинки, у хитонов метамерии группируются и мышцы тела и кровеносные сосуды; другими словами, мезодермальиые полоски их получают сегментировку, зависящую от метамерии эктодермальных образований, т. е. такую же. какая возникает в мезодермальных полосках ларвальных сегментов аннелид.
У Aplacophora (Neomeria) трохофора характеризуется крупной величиной клеток и величиной прототроха, надвигающегося на посторальную половину тела и совершеннозакрывающего ее (рис. 106). При метаморфозе эта половина растет в длину, а прототрох отстает в развитии и исчезает; на боках этой половины появляются пластинки (7), число которых впоследствии увеличивается, но они приобретают уже форму иголочек.  Пластинки сближают молодых Aplacophora с хитонами, но, с другой стороны, и у хитонов при метаморфозе появляется боковой ряд иголочек. Метаморфоз типичной трохофоры боконервных моллюсков в общем отличается от того типа метаморфоза, который описан в начале этой главы, как характерный для большинства моллюсков. У хитонов метаморфоз трохофоры происходит путем почти равномерного разрастания всего тела с некоторым преобладанием роста в длину. Вырастание из производных 2 d мантии и ноги мало отличается по интенсивности от остальных частей тела. Поэтому и нога и мантиялишь немного выступают наружу, и довольно крупные размеры их являются результатом роста всего тела в длину иотчасти в ширину. С удлинением тела и мантии удлиняется и зачаток раковины, а раковина, покрывающая тело во всю его длину, не стесняет сгибания тела, необходимого для этих моллюсков, только в том случае, если она будет расчленена на отдельные метамерные пластинки; это и имеет место у хитонов. Метамерное же расчленение раковины вызывает и частичную метамерию мышц и некоторых других мезодермальных производных. Рост при метаморфозе в длину сближает морфологию тела хитонов с морфологией ларвальных сегментов аннелид, и это сходство увеличивается частичной метамерией тела, зависящей от расчленения раковины так же, как у аннелид ларвальная метамерия зависит от сегментированного расположения органов движения — параподий.
Пластинки сближают молодых Aplacophora с хитонами, но, с другой стороны, и у хитонов при метаморфозе появляется боковой ряд иголочек. Метаморфоз типичной трохофоры боконервных моллюсков в общем отличается от того типа метаморфоза, который описан в начале этой главы, как характерный для большинства моллюсков. У хитонов метаморфоз трохофоры происходит путем почти равномерного разрастания всего тела с некоторым преобладанием роста в длину. Вырастание из производных 2 d мантии и ноги мало отличается по интенсивности от остальных частей тела. Поэтому и нога и мантиялишь немного выступают наружу, и довольно крупные размеры их являются результатом роста всего тела в длину иотчасти в ширину. С удлинением тела и мантии удлиняется и зачаток раковины, а раковина, покрывающая тело во всю его длину, не стесняет сгибания тела, необходимого для этих моллюсков, только в том случае, если она будет расчленена на отдельные метамерные пластинки; это и имеет место у хитонов. Метамерное же расчленение раковины вызывает и частичную метамерию мышц и некоторых других мезодермальных производных. Рост при метаморфозе в длину сближает морфологию тела хитонов с морфологией ларвальных сегментов аннелид, и это сходство увеличивается частичной метамерией тела, зависящей от расчленения раковины так же, как у аннелид ларвальная метамерия зависит от сегментированного расположения органов движения — параподий.
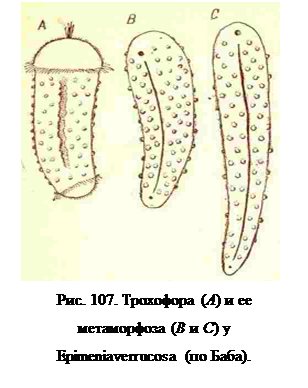 Еще больше от типичного метаморфоза моллюсков отличается метаморфоз Aplacophora. (Epimenia). Рост тела трохофоры в длину у них настолько преобладает над ростом в других направлениях, что тело моллюска становится червевидным; при этом цилиндрическая форма тела трохофоры сохраняется и у взрослого моллюска. Производные 2 d образуют дорзальную и боковые стенки червовидного тела и не дают своим ростом ни мантии, ни ног. Отсутствие закладки этих типичных для моллюсков органов в онтогенезе Aplacophoraнесомненно является доказательством примитивности всех особенностей строения этих червовидных моллюсков. Так как нога в большинстве других классов моллюсков возникает из краев бластопора, составляющих в то же время и вентральные края соматобласта (производных 2 d), а при своем развитии пора закрывает весь бластопор, кроме переднего и заднего его концов, то отсутствие зачатка ноги у Aplacophora определяет и очень своеобразную у них бластопора. Он также закрывается в средней своей части, но закрывается, как и у аннелид, только производными третьего квартета; образуемая здесь этим последним тонкая эпителиальная пластинка образует на месте бластопора узкий желобок, на переднем конце которого находится ротовое, на заднем — анальное отверстие. Так как при метаморфозе растет в длину все тело трохофоры, то соответственно сильно удлиняется бластопор или желобок, соответствующий ему и превращающийся в продольный желоб на вентральной стороне тела взрослого моллюска.
Еще больше от типичного метаморфоза моллюсков отличается метаморфоз Aplacophora. (Epimenia). Рост тела трохофоры в длину у них настолько преобладает над ростом в других направлениях, что тело моллюска становится червевидным; при этом цилиндрическая форма тела трохофоры сохраняется и у взрослого моллюска. Производные 2 d образуют дорзальную и боковые стенки червовидного тела и не дают своим ростом ни мантии, ни ног. Отсутствие закладки этих типичных для моллюсков органов в онтогенезе Aplacophoraнесомненно является доказательством примитивности всех особенностей строения этих червовидных моллюсков. Так как нога в большинстве других классов моллюсков возникает из краев бластопора, составляющих в то же время и вентральные края соматобласта (производных 2 d), а при своем развитии пора закрывает весь бластопор, кроме переднего и заднего его концов, то отсутствие зачатка ноги у Aplacophora определяет и очень своеобразную у них бластопора. Он также закрывается в средней своей части, но закрывается, как и у аннелид, только производными третьего квартета; образуемая здесь этим последним тонкая эпителиальная пластинка образует на месте бластопора узкий желобок, на переднем конце которого находится ротовое, на заднем — анальное отверстие. Так как при метаморфозе растет в длину все тело трохофоры, то соответственно сильно удлиняется бластопор или желобок, соответствующий ему и превращающийся в продольный желоб на вентральной стороне тела взрослого моллюска.
Брюхоногие (Gastropoda)
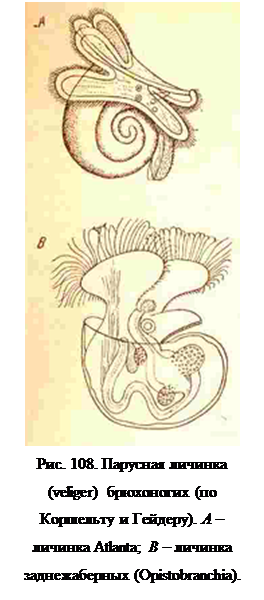
Наиболее примитивные в морфологическом отношении представители переднежаберных (Prosobranchia), как Fissurella, Haliotis, Patella, Acmaea, выходит из яйца в виде типичной трохофоры, похожей на трохофору аннелид (рис. 77). По внутреннему строению она отличается от последней слабым развитием мезенхимы, отсутствием протонефридиев и ранним разрыхлением мезодермальных полосок, которые вскоре во всех своих частях, кроме заднего участка, распадаются на рассеивающиеся в полости тела клетки. У остальных брюхоногих, так же как и у большинства других моллюсков, личинкой бывает veliger (парусник, рис. 108). Он имеет зачаток ноги на брюшной и раковинную железу на спинной стороне, причем последняя уже вывернута наружу и имеет очень тонкую раковину. Эти образования увеличивают вес личинки, но она не надает на дно, как личинка хитона, а продолжает плавать, благодаря ресничкам паруса.
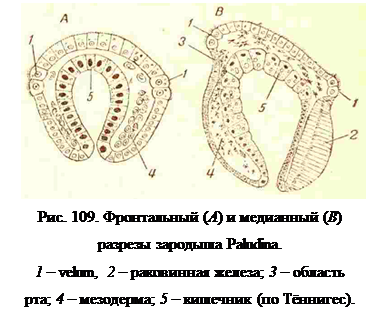
Если из яйца выходит veliger. то предшествующая ему стадия трохофоры все-таки может плавать внутри яйцевой оболочки при помощи своего прототроха и в таком плавающем состоянии существует даже у пресноводных живородящих переднежаберных, как, например, у Paludina, в качестве одной из ранних эмбриональных стадий (рис. 109); из яйца Paludina выходит в виде уже почти готового моллюска.
У морских брюхоногих из яйца всегда выходит личинка, но у различных видов степень развития veliger бывает различна, в зависимости от количества желтка в яйце. Veliger может быть снабжен более или менее развитой мантией и раковинкой, а его внутренние органы могут находиться у одних моллюском на более ранних, у других на более поздних стадиях развития. В случаях далеко продвинувшейся внутренней организация у личинки могут возникать специальные провизорные органы выделения, однако, вес-таки не в виде протонефридиев, а в виде крупных клеток, прилегающих изнутри к эктодерме в передней части тела (рис. 110).
 Количество желтка в яйце, конечно, оказывает влияние на дробление, которое и в этих случаях остается спиральным, с хорошо выраженным апикальным крестом, но бластомеры основного квартета по объему в несколько сот раз превосходят бластомерыдругих квартетов, которые собраны на анимальном полюсе в виде сравнительно маленькой пластинки (рис. 111). При образовании четвертого квартета бластомеры 4 а, 4 b и 4 c отделяются с большим количеством желтка в виде очень крупных бластомер, а 4 d не получает желтка и несравненно меньше их по размерам.
Количество желтка в яйце, конечно, оказывает влияние на дробление, которое и в этих случаях остается спиральным, с хорошо выраженным апикальным крестом, но бластомеры основного квартета по объему в несколько сот раз превосходят бластомерыдругих квартетов, которые собраны на анимальном полюсе в виде сравнительно маленькой пластинки (рис. 111). При образовании четвертого квартета бластомеры 4 а, 4 b и 4 c отделяются с большим количеством желтка в виде очень крупных бластомер, а 4 d не получает желтка и несравненно меньше их по размерам.
У многих брюхоногих и у Dentalium (рис. 78) дробление сопровождается образованием полярной лопасти, подобной полярной лопасти Chaetopterus и Myzostomum среди аннелид. Несмотря на то, что полярная лопасть связана с квадрантом D, размеры 2 d и 4 d бывают в таких случаях лишь немного крупнее размеров других бластомер тех же квадрантов.
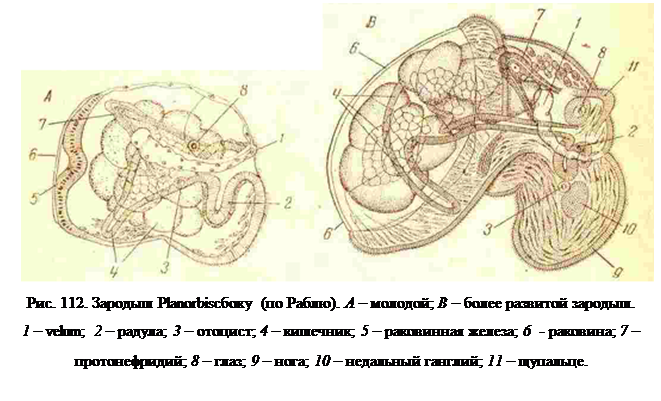
 У пресноводных и наземных легочных моллюсков (Pulmonata) яйцо развивается в коконе, окружено в нем белком и имеет очень мелкие размеры, так как содержит очень мало желтка. Принимая во внимание то, что известно о влиянии питания белком на развитие некоторых червей, можно было и здесь ожидать того же. Однако, у легочных (Pulmonata), так же как у олигохет и пиявок, белок не влияет на дробление, которое целиком проходит и яйце, одетом собственной первичной оболочкой. Последняя сбрасывается при начале формирования зародыша, и только тогда белок получает к нему доступ. Поэтому дробление Pulmonata очень хорошо выраженное спиральное, большей частью без преобладания размеров квадранта D, и из первичной яйцевой оболочки в белок выходит округлый зародыш с признаками veliger. Лопасти паруса (velum) или разросшегося прототроха ясно выражены, но невелики, и зародыш активно плавает (вращается) в белке при помощи мерцательных волосков, одевающих всю его голову и зачаток ноги (рис. 112). У зародыша легочников имеется и слабо выраженная характерная для трохофоры теменная пластинка, есть уже раковинная железа с нежной маленькой раковинкой, а также отоцисты и радула. Так как в кокон проникает извне пресная вода, и возникает вследствие, этого потребность в осморегуляции, то у таких эмбрионов-личинок возникают протонефридии (рис. 112)как осморегулирующий орган, отсутствующий у морских личинок моллюсков.
У пресноводных и наземных легочных моллюсков (Pulmonata) яйцо развивается в коконе, окружено в нем белком и имеет очень мелкие размеры, так как содержит очень мало желтка. Принимая во внимание то, что известно о влиянии питания белком на развитие некоторых червей, можно было и здесь ожидать того же. Однако, у легочных (Pulmonata), так же как у олигохет и пиявок, белок не влияет на дробление, которое целиком проходит и яйце, одетом собственной первичной оболочкой. Последняя сбрасывается при начале формирования зародыша, и только тогда белок получает к нему доступ. Поэтому дробление Pulmonata очень хорошо выраженное спиральное, большей частью без преобладания размеров квадранта D, и из первичной яйцевой оболочки в белок выходит округлый зародыш с признаками veliger. Лопасти паруса (velum) или разросшегося прототроха ясно выражены, но невелики, и зародыш активно плавает (вращается) в белке при помощи мерцательных волосков, одевающих всю его голову и зачаток ноги (рис. 112). У зародыша легочников имеется и слабо выраженная характерная для трохофоры теменная пластинка, есть уже раковинная железа с нежной маленькой раковинкой, а также отоцисты и радула. Так как в кокон проникает извне пресная вода, и возникает вследствие, этого потребность в осморегуляции, то у таких эмбрионов-личинок возникают протонефридии (рис. 112)как осморегулирующий орган, отсутствующий у морских личинок моллюсков.
 Органотипический период развития меньше подвергается изменяющему действию белка как среды; но у сухопутных Pulmonata (Helix. Limax, Arion), в кокон которых проникает дождевая вода, более поздние стадии развития могут сильно изменяться, повидимому, вследствие низкого осмотического давления среды. Зародыш сильно раздувается в передней своей части (рис. 113), образуя головной пузырь, по объему превышающий все остальное тело зародыша; несколько меньшей величины пузырь вздувается в ноге (так называемый подоцист). Необходимая циркуляция переполняющей тело жидкости обеспечивается, или пульсацией всех стенок
Органотипический период развития меньше подвергается изменяющему действию белка как среды; но у сухопутных Pulmonata (Helix. Limax, Arion), в кокон которых проникает дождевая вода, более поздние стадии развития могут сильно изменяться, повидимому, вследствие низкого осмотического давления среды. Зародыш сильно раздувается в передней своей части (рис. 113), образуя головной пузырь, по объему превышающий все остальное тело зародыша; несколько меньшей величины пузырь вздувается в ноге (так называемый подоцист). Необходимая циркуляция переполняющей тело жидкости обеспечивается, или пульсацией всех стенок
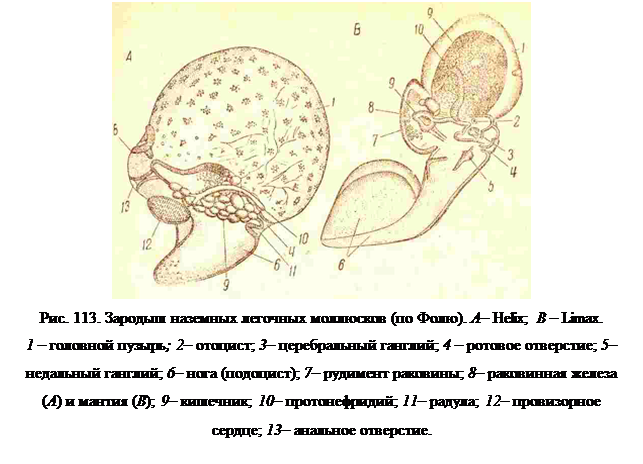 Пузыря, или пульсацией сердца. Последнее представляет собой выпяченный наружу эктодермальный карман (рис. 113, А, 12), содержащий некоторое количество функционирующих у зародыша мускульных клеток, и не имеет никакого отношения к сердцу взрослого животного, возникающего вместе с перикардием из мезодермы. Личиночное сердце встречается, однако, и у личинок некоторых морских переднежаберников.
Пузыря, или пульсацией сердца. Последнее представляет собой выпяченный наружу эктодермальный карман (рис. 113, А, 12), содержащий некоторое количество функционирующих у зародыша мускульных клеток, и не имеет никакого отношения к сердцу взрослого животного, возникающего вместе с перикардием из мезодермы. Личиночное сердце встречается, однако, и у личинок некоторых морских переднежаберников.
Характерными особенностями большинства брюхоногих являются несимметричное строение тела и спирально закрученная раковина, представляющая собою как бы слепок со спирально закрученного мешка мантии, в который перемещается большая часть внутренних органов. Обе эти особенности появляются уже во время метаморфоза, так как и трохофора и veliger вполне билатерально симметричны: анальное отверстие лежит на одной продольной линии с ротовым, нога вырастает по середине вентральной стороны, раковина же и мантия находятся по срединной линии спинной стороны, причем раковина или имеет вид выпуклого диска,, или загнута  своей вершиной к голове в сагиттальной плоскости (рис. 114, А).
своей вершиной к голове в сагиттальной плоскости (рис. 114, А).
Еще во время плавания личинки в течение сравнительно небольшого промежутка времени раковина целиком начинает поворачиваться вместе с мантией вправо и описывает своей загнутой вершиной дугу в 1800, так что вершина ее оборачивается назад (рис. 114, В). Этот поворот раковины происходит вследствие того, что спинная часть тела veliger, с которой соединена мантия, вступает в период усиленного роста, но растетпри этом только левая сторона спины, правая же или не растет или растет очень медленно. Вследствие этого анальное отверстие и все образующиеся около него органы (жабры, сердце, почки) перемещаются вперед и на правую сторону. Кроме того, мантийная полость, появившаяся в виде вдающейся складки под краем мантии над анальным отверстием, также растет несимметрично и углубляется главным образом в правойсвоей половине. Совместное действие роста левой стороны спины в горизонтальном круговом направлении и правой стороны вглубь полости мантии, т. е. кверху, приводит к тому, что анальное отверстие, во-первых, ложится на более высоком уровне, чем ротовое, и во-вторых, оно вместе с раковиной делает поворот на 180°, так что опять располагается на одной продольной линии с ротовым отверстием, но не на противоположномконце тела, а почти прямо над ртом.
 Такое положение занимают анальное отверстие и прилегающие к нему органы так называемого мантийного комплекса только у примитивных представителей переднежаберников, у остальных же, а также у легочных и заднежаберных анальное отверстие описывает в большей или меньшей степени неполную дугу и останавливается па правой стороне тела, причем из сопровождающих анальное отверстие парных органов, т. е. почек, жабер, предсердий сердца, развиваются только левые.
Такое положение занимают анальное отверстие и прилегающие к нему органы так называемого мантийного комплекса только у примитивных представителей переднежаберников, у остальных же, а также у легочных и заднежаберных анальное отверстие описывает в большей или меньшей степени неполную дугу и останавливается па правой стороне тела, причем из сопровождающих анальное отверстие парных органов, т. е. почек, жабер, предсердий сердца, развиваются только левые.
Спиральное закручивание мантийного мешка и раковины наступает значительно позднее, чем только что описанное поворачивание мантийного мешка на его основании; оно происходит во время превращения veliger в моллюска. В это время поверхность мантии перестает вырабатывать раковину, которая продолжает образовываться только деятельностью края мантии. Мантийный мешок при этом растет сильно в вышину, но рост его несимметричен, обычно правая сторона растет медленнее левой, хотя бывает и наоборот — правая растет быстрее левой. Вследствие неровного роста, та сторона конического мантийного мешка, которая растет быстрее, обвивается вокруг более медленно растущей стороны.
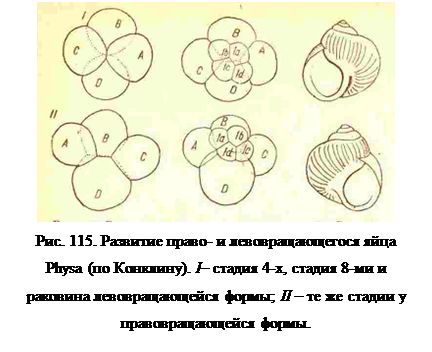
 Правозакрученная и левозакрученная раковины встречаются даже у одного и того же вида брюхоногих, как наследственные мутации. У этих же видов брюхоногих выделение квартетов обычноначинается дексиотропно, т. е. третье и последующие нечетные деления сопровождаются дексиотропным смещением анимальной протоплазмы и анимальных бластомер, а четные деления происходят леотропно; но отделение квартетов может начинаться у некоторых особей того же вида леотропно. Из яйца с дексиотропньм началом выделения квартетов развивается моллюск с правозакрученной раковиной, а из яйца с леотропным отделением первого квартета моллюск получается с лепозакрученной раковиной (рис. 115). Кроме того установлено, что дексиотропно дробятся те яйца, которые, будучи ооцитами в половой железе, прикреплены вегетативным полюсом к стенке железы, а леотропно начинают дробиться яйца, прикрепленные анимальным полюсом к стенке половой железы. Все эти три момента, т. е. способ прикрепления ооцита, направление смещения бластомер при первом и третьем делении и направление закручивания раковины, являются признаками одной и той же наследственной мутации. Но связь этих моментов между собою представляет большой теоретический интерес, так как показывает, что предварительная структура яйца, частично зависящая от положения ооцита в яичнике, определяет не только характер его дробления, но и некоторые особенности взрослого организма через данный характер дробления, т. е. дексиотропная тенденция начала дробления выражается у взрослого организма также и в закручивании раковины по тому леже правому направлению.
Правозакрученная и левозакрученная раковины встречаются даже у одного и того же вида брюхоногих, как наследственные мутации. У этих же видов брюхоногих выделение квартетов обычноначинается дексиотропно, т. е. третье и последующие нечетные деления сопровождаются дексиотропным смещением анимальной протоплазмы и анимальных бластомер, а четные деления происходят леотропно; но отделение квартетов может начинаться у некоторых особей того же вида леотропно. Из яйца с дексиотропньм началом выделения квартетов развивается моллюск с правозакрученной раковиной, а из яйца с леотропным отделением первого квартета моллюск получается с лепозакрученной раковиной (рис. 115). Кроме того установлено, что дексиотропно дробятся те яйца, которые, будучи ооцитами в половой железе, прикреплены вегетативным полюсом к стенке железы, а леотропно начинают дробиться яйца, прикрепленные анимальным полюсом к стенке половой железы. Все эти три момента, т. е. способ прикрепления ооцита, направление смещения бластомер при первом и третьем делении и направление закручивания раковины, являются признаками одной и той же наследственной мутации. Но связь этих моментов между собою представляет большой теоретический интерес, так как показывает, что предварительная структура яйца, частично зависящая от положения ооцита в яичнике, определяет не только характер его дробления, но и некоторые особенности взрослого организма через данный характер дробления, т. е. дексиотропная тенденция начала дробления выражается у взрослого организма также и в закручивании раковины по тому леже правому направлению.