По сравнению с яйцами остальных моллюсков (0,03—0,4 мм) яйца головоногих имеют громадные размеры (10-15 мм), так как в изобилии содержат желток, который составляет главную массу яйца. Протоплазма яйца в виде тонкого слоя занимает приблизительно  поверхности яйца, так что объем ее приблизительно в 400 раз меньше объема желтка. Яйцо имеет почти эллипсоидальную форму, причем одна из длинных сторон его несколько сплюснута (рис. 123). Протоплазма занимает область вокруг одного из полюсов яйца, и эта область отличается от желтка более светлой окраской. Положение развивающегося зародыша детерминировано и по отношению к сторонам будущего животного: анимальное полушарие яйца до дробления является дорзальным, вегетативное — вентральным, сплюснутая длинная сторона — задней стороной, а выпуклая, противоположная ей — передней стороной (рис. 123). Яйцо относится телолецитальному типу, дробление идет дискоидально и не имеет никаких признаков спирального дробления, свойственного всем остальным моллюскам. Делится только протоплазма зародышевого диска, в желток же борозды дробления проникают на незначительную глубинутолько во время дробления, а затем все эмбриональные процессы идут в клетках диска, желток же остается пассивным.
поверхности яйца, так что объем ее приблизительно в 400 раз меньше объема желтка. Яйцо имеет почти эллипсоидальную форму, причем одна из длинных сторон его несколько сплюснута (рис. 123). Протоплазма занимает область вокруг одного из полюсов яйца, и эта область отличается от желтка более светлой окраской. Положение развивающегося зародыша детерминировано и по отношению к сторонам будущего животного: анимальное полушарие яйца до дробления является дорзальным, вегетативное — вентральным, сплюснутая длинная сторона — задней стороной, а выпуклая, противоположная ей — передней стороной (рис. 123). Яйцо относится телолецитальному типу, дробление идет дискоидально и не имеет никаких признаков спирального дробления, свойственного всем остальным моллюскам. Делится только протоплазма зародышевого диска, в желток же борозды дробления проникают на незначительную глубинутолько во время дробления, а затем все эмбриональные процессы идут в клетках диска, желток же остается пассивным.

 Первая борозда дробления делит зародышевый диск в направлении сагиттальной плоскости тела будущего зародыша, т. е. по линии, идущей по меридиану от наиболее выпуклой длинной стороны к наименее выпуклой (рис. 124). Вторая борозда также проходит меридианально, перпендикулярно к плоскости первой борозды, причем из получающихся четырех бластомер две передние крупнее двух задних (рис. 124, В и 125, А).
Первая борозда дробления делит зародышевый диск в направлении сагиттальной плоскости тела будущего зародыша, т. е. по линии, идущей по меридиану от наиболее выпуклой длинной стороны к наименее выпуклой (рис. 124). Вторая борозда также проходит меридианально, перпендикулярно к плоскости первой борозды, причем из получающихся четырех бластомер две передние крупнее двух задних (рис. 124, В и 125, А).
Также меридианально проходят и две третьих борозды, причем они разделяют каждую из двух передних бластомер на две равные половины, а каждую из двух задних делят так, что, проходя параллельно первой борозде и близко к ней, отделяют две узенькие бластомеры, лежащие в задней половине по бокам медиальной плоскости (рис. 124, В и 125, А).
Толщина слоя протоплазмы зародышевого диска настолько незначительна, что даже и после третьего и четвертого деления протяженность его протоплазмы остается наибольшей в направлении, параллельном поверхности яйца. В этом же направлении ложатся и веретена четвертого и пятого делений, которые также разделяют диск в меридианальном направлении. При этом уже со второго деления группировка бластомер свидетельствует о билатеральности дробления (рис. 124 и 125), да и яйцо до дробления имеет форму, в которой может быть только одна плоскость симметрии.
 Меридианальные борозды у головоногих очень мало проникают в желток, и бластомеры первых четырех делений остаются соединенными с желтком своей периферической частью; при пятом же делении, направленные к центру концы некоторых бластомер отделяются в виде свободных клеток от периферических частей этих бластомер, сохраняющих соединение с желтком. Тоже происходит и при последующих делениях дробления, так что центральное поле диска состоит на этих стадиях из постепенно увеличивающегося количества свободных мелких клеток, а периферия диска состоит из бластомер, соединенных е желтком и получивших название бластоконов (рис. 126, А). Увеличение числа клеток центрального поля происходит частью поперечным делением бластоконов, частью размножением самих свободных бластомер, и вследствие этого диск постепенно расширяется по всем направлениям, а бластоконы смещаются все дальше от полюса, сохраняя свое положение на периферии диска.
Меридианальные борозды у головоногих очень мало проникают в желток, и бластомеры первых четырех делений остаются соединенными с желтком своей периферической частью; при пятом же делении, направленные к центру концы некоторых бластомер отделяются в виде свободных клеток от периферических частей этих бластомер, сохраняющих соединение с желтком. Тоже происходит и при последующих делениях дробления, так что центральное поле диска состоит на этих стадиях из постепенно увеличивающегося количества свободных мелких клеток, а периферия диска состоит из бластомер, соединенных е желтком и получивших название бластоконов (рис. 126, А). Увеличение числа клеток центрального поля происходит частью поперечным делением бластоконов, частью размножением самих свободных бластомер, и вследствие этого диск постепенно расширяется по всем направлениям, а бластоконы смещаются все дальше от полюса, сохраняя свое положение на периферии диска.
 Отделение новых бластомер от бластоконов вскоре прекращается, и дальнейший рост диска обусловливается исключительно делением свободных клеток; с этого момента несколько расплывчатые очертания диска становятся гораздо более отчетливыми, а бластоконы обособляются от его краев и делением образуют группы клеток на желткеу краев диска (рис. 126, В). Из этих клеточных групп па поверхности желтка образуется сплошной эпителиальный слой, состоящий из сильно распластанных на желтке клеток. Его называют желточным эпителием,или желточной энтодермой.
Отделение новых бластомер от бластоконов вскоре прекращается, и дальнейший рост диска обусловливается исключительно делением свободных клеток; с этого момента несколько расплывчатые очертания диска становятся гораздо более отчетливыми, а бластоконы обособляются от его краев и делением образуют группы клеток на желткеу краев диска (рис. 126, В). Из этих клеточных групп па поверхности желтка образуется сплошной эпителиальный слой, состоящий из сильно распластанных на желтке клеток. Его называют желточным эпителием,или желточной энтодермой.
По мере центробежного надвигания клеток зародышевого диска на желток, края диска, вследствие делямнинации или подворачивания края, становятся многослойными и выступают над поверхностью желтка. Клетки нижнего слоя этого краевого утолщения распространяются подо всей поверхностью бластодиска в видекишечной энтодермы. Также из краевого утолщения, по некоторым данным независимо от энтодермы и позднее закладки последней, появляется мезодерма в виде группы клеток, разрастающихся и проникающих между эктодермой (наружным слоем бластодиска) и кишечнойэнтодермой. Несколько позже появляется невыясненным способом группа клеток, которые долго остаются пассивными  и представляют собою половой зачаток.
и представляют собою половой зачаток.
После образования зародышевых листков мезодерма и эктодерма начинают быстро разрастаться непрерывным слоем по поверхности желточной энтодермы. В это время разрастаются также эктодерма и мезодерма в самом бластодиске, образуя в нем утолщения, складки п выросты, связанные с процессом формообразования.
Образование внешней формы зародыша начинается появлением в центре бластодиска, т. е.в анимальной или дорзальной части зародыша, небольшого бугорка, верх ушка которого образует впячение—зачаток раковинной железы (рис. 127, А).
На несколько более поздних стадиях края этого впячения сближаются и срастаются так, та к что раковинная железа образует замкнутый мешочек и в виде мошка разрастается по дорзальной стороне по мере вырастания зародыша. Вслед за появлением раковинной железы на правой и левой частях бластодиска образуются вздутия вследствие разрастания эктодермы. Эти вздутия представляют собою зачатки глаз и оптических ганглиев, причем сами глаза появляются на дорзальной стороне этих вздутий в виде впячений или глазных ямок, замыкающихся затем в глазные пузыри (рис. 127, В и С). На дорзальной же стороне между глазными ямками возникает в виде впячения стомодеум, а у основания бугорка раковинной железы закладывается небольшой кольцевой валик — зачаток мантии (рис. 128, 2).
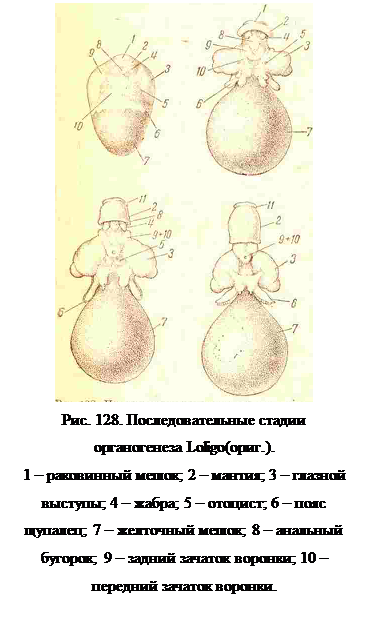 Еще несколько позже раковинная железа приподнимается над поверхностью зародыша на вершине бугорка, представляющего собою зачаток туловища моллюска. Между ним и глазными зачатками возникает еще кольцевой валик — зачаток воронки с парой отоцистов у его основания, а на самом зачатке туловища, на вентральной стороне, появляется пара удлиненных бугорков — зачатки жабер, а между ними, по срединной линии. — впячение проктодеума (рис. 127, С, 6). На границе зародыша и желточного мешка закладываются щупальца в виде кольцевого ряда бугорков (рис- 128,6).
Еще несколько позже раковинная железа приподнимается над поверхностью зародыша на вершине бугорка, представляющего собою зачаток туловища моллюска. Между ним и глазными зачатками возникает еще кольцевой валик — зачаток воронки с парой отоцистов у его основания, а на самом зачатке туловища, на вентральной стороне, появляется пара удлиненных бугорков — зачатки жабер, а между ними, по срединной линии. — впячение проктодеума (рис. 127, С, 6). На границе зародыша и желточного мешка закладываются щупальца в виде кольцевого ряда бугорков (рис- 128,6).
На следующих стадиях мантия нарастает на туловищный бугорок и прикрывает сначала раковинный мешочек, а затем зачатки жабер и проктодеум; на мантии появляются при этом боковые плавнички. Зачатки щупалец перемещаются к дорзальной стороне головы и окружают стебелек, которым зародыш соединен с желточным мешком (рис. 128, 6). По мере закладки и разрастания всех этих зачатков, последние надавливают на желток, который принимает в области туловищного зачатка коническую суженную форму, в то время как внезародышевый желток остается грушевидным. На месте соединения зародышевой и внезародышевой половин желтка образуется резкий перехват. Потребление желтка идет наиболее интенсивно в задней части зародыша,и на место желтка распространяется мезодермальная ткань, из которой здесь формируется перикардий и сердце (рис. 129).
Мы видели, что эпителий, непосредственно одевающий желточный меток, возникает из бластоконов, как желточная энтодерма; внутри самого зародыша желток одет кишечным эпителием, возникающим из краевого утолщения бластодиска. В одном небольшом участке этой кишечной энтодермы клетки собраны плотной группой, которая на более поздних стадиях отщепляется от эпителия желтка в виде плотного эпителиального мешочка. Этот мешочек представляет собою зачаток средней кишки, который затем растет в виде трубки по направлению к проктодеуму, другим же концом упирается в желток. После потребления той части желтка, которая находится на уровне анального отверстия, конец зачатка средней кишки растет несколько дальше и образует расширение, из которого формируется желудок (рис. 129, В, 17) и, кроме того, в виде выростов его возникают дольки печени. В то же время от стомодеума растет к этому расширениюпрямая трубка — зачаток пищевода, который соединяется со средней кишкой (рис. 129, В, 15). Таким образом, не только энтодерма желточного метка, но и большая часть энтодермы, одевающей желток в зародыше, служит только для усвоения зародышем желтка и никакого участия в образовании кишечника не принимает; последний образуется из небольшого участка энтодермы зародыша и начинает развиваться только после того, как обособится совсем от желтка.
 Желточный мешок занимает на теле зародыша головоногих положение, какое не встречается больше ни у каких животных. У большинства беспозвоночных животных желток занимает спинную сторону зародыша, у позвоночных — брюшную, у головоногих же желток лежит на голове, как бы выпячивается изо рта зародыша. На самом деле, однако, рот лежит несколько дорзально от желточного стебелька (рис. 129, В, 12) и только после исчезновения желтка становится на его место — в центре венца щупалец.
Желточный мешок занимает на теле зародыша головоногих положение, какое не встречается больше ни у каких животных. У большинства беспозвоночных животных желток занимает спинную сторону зародыша, у позвоночных — брюшную, у головоногих же желток лежит на голове, как бы выпячивается изо рта зародыша. На самом деле, однако, рот лежит несколько дорзально от желточного стебелька (рис. 129, В, 12) и только после исчезновения желтка становится на его место — в центре венца щупалец.
Сначала зародыш по объему меньше желточного мешка, но он растет но мере потребления желтка, и отношения объемов становятся обратными.
Как видно из всего сказанного о развитии моллюсков, головоногие стоят в этом отношении совершенно особняком, хотя органогенез и идет по общему типу органогенеза моллюсков, т. е. закладываются в определенных топографических взаимоотношениях мантия, раковина, голова и нога; впрочем последняя очень своеобразно разделена, на воронку с отоцистами и щупальца. Поэтому не может быть никаких сомнений в принадлежности головоногих к моллюскам, и возникает вопрос, от чего зависят те отклонения в ранних стадиях развития головоногих, которые делают их онтогенез совершенно несравнимым со спиральным дроблением других моллюсков.
Головоногие являются наиболее высоко организованным классом моллюсков, так как все части и все органы достигают у них наиболее полного и сложного развития. Судя по отсутствию каких-либо следов метаморфоза, головоногие начали очень давно эволюционировать с неизменной установкой на взрослую форму. Это видно из того, чтоне только расположение бластомер, но и самое строение яйца до начала развития имеет отчетливый билатерально симметричный характер, а оси и плоскость симметрии яйца отвечают осям и плоскости симметрии взрослого животного. В этом лежит первая причина отсутствия у них каких-либо признаков спирального дробления, ведущего к образованию по внешности радиально симметричной трохофоры. Второй же причиной является то, что яйцо для развития до взрослой формы, имеющей притом сложное и специализированное строение, должно быть снабжено очень большим количеством желтка. Поэтому дробление у них идет по дискоидальному типу и сопровождается образованием специальной, категории клеток, — желточной энтодермы, предназначенной для переработки всей массы желтка во время развития остальных органов животного, включая сюда и дефинитивный кишечник.