 Дробление различных Entomostraca сильно варьирует как вследствие различного содержания в яйцах желтка, так и в силу неодинакового порядка и способа закладки первичных зачатков.
Дробление различных Entomostraca сильно варьирует как вследствие различного содержания в яйцах желтка, так и в силу неодинакового порядка и способа закладки первичных зачатков.
Яйца с наименьшим количеством желтка свойственны свободно живущим веслоногим (Copepoda) и некоторым из ветвистоусых (Cladocеra), развивающихся из летних, партеногенетических яиц. Дробление яйца циклопа (Copepoda) и Polyphemus (Cladocеra) — полное; первые две борозды, как обычно, меридианальные, третья — экваториальная, причем бластомеры анимального полушария ложатся над промежутками бластомер вегетативного (рис. 130, В), что напоминает расположение бластомер на этой стадии при спиральном дроблении. Бластомеры вегетативного полушария при следующем делении ложатся иначе, чем бластомеры анимального. Если бластомеры стадии 4 бластомер обозначить буквами A, B, C иD то на стадии 8 бластомер все бластомеры анимального полушария (a, b, c иd) детерминированы на образование так называемой первичной эктодермы. У Polyphemus в вегетативном полушарии бластомера Dна стадии 32 бластомер разделяется на четыре (D11,D12,D21 и D22); одна из них (D11) представляет собою первичную половую клетку, другая (D12) — первичную энтодермальную (рис. 132, В и С); остальные две вместе с прочими бластомерами вегетативного полушария окружают венцом половую и энтодермальную бластомеры (рис. 132, D). На стадии 64 клеток бластомеры этого круга разделяются на два венца: внутренний, непосредственно прилегающий к половой и энтодермальной бластомерам, и наружный; внутренний венец является мезодермой, а наружный — вторичной эктодермой. Таким образом,  бластомер D дает эктодерму, энтодерму, мезодерму и половой зачаток, а остальные вегетативные бластомеры дают только вторичную эктодерму и мезодерму. После обособления всех этих зачатков детерминированность дробления исчезает и клетка мезодермальная может стать эктодермальной и наоборот.
бластомер D дает эктодерму, энтодерму, мезодерму и половой зачаток, а остальные вегетативные бластомеры дают только вторичную эктодерму и мезодерму. После обособления всех этих зачатков детерминированность дробления исчезает и клетка мезодермальная может стать эктодермальной и наоборот.
Тот путь, которым идет обособление первичной половой клетки, иначе говоря, зачатковый путь у циклопа очень ясно прослеживается, благодаря тому, что при первом жеделении яйца на одном конце веретена деления, лежащем в СD, появляются в протоплазме зернышки, так называемые эктозомы (рис. 131, 1), которые имеют плазменноепроисхождение, т. е., в отличие от отбрасываемых в протоплазму частиц у Ascaris, состоят не из хроматина. Эктозомы растворяются в протоплазме, но на конце веретена второго деления, лежащем в блаетомере D, они снова появляются, и так продолжается до пятого деления включительно, когда они обнаруживаются в первичной половой клетке.
 При дроблении летних яиц Polyphemus (Cladoсera, рис. 132), детерминированность сохраняется дольше, чем у циклопа, и можно проследить судьбу частипервичной эктодермы вплоть до образования из нее теменной пластинки (рис. 133).
При дроблении летних яиц Polyphemus (Cladoсera, рис. 132), детерминированность сохраняется дольше, чем у циклопа, и можно проследить судьбу частипервичной эктодермы вплоть до образования из нее теменной пластинки (рис. 133).
У Lepas (уеоиогие — Cirripеdia,) яйцо содержит значительно больше желтка, который при первом делении попадает весь вбластомеру СD, а при втором — в D (рис. 134). АВ дает эктодерму, а Dделится только один раз, отделяя от себя богатую протоплазмой первичную мезодермальную клетку, а сама становится клеткой энтодермы. В этом случае мы опять встречаем связь мезодермы с энтодермой, как при спиральном дроблении, связь, которая находит себе объяснении в том, что весь питательный материал находится в D. Небольшая часть мезодермы возникает, впрочем, из эктодермальных клеток, что напоминает эктомезодерму трохофоры, но едва ли они вполне гомологичны друг другу.

 При увеличении количества желтка в яйце дробление Eutomostraca меняет свой характер. Так, в яйцах ракушковых (Ostracoda. рис. 135) и в зимних яйцах Daphina дробление остается полным до гаструляции, но после гаструляции (путем иммиграции) внутренние клетки сливаются, образуя желточную внутреннюю массу с ядрами. У Ostracoda ядра, в желтке, располагаются при этом на правильных расстояниях друг от друга, так что возможно, что полного исчезновения границ клеток и не происходит, но в зимних яйцах Daphina ядра во внутренней желточной массе рассеяны беспорядочно, чтоговорит о несомненном слиянии клеток. Клетки наружного слоя — эктодермы — не сливаются друг с другом, образуя наружный эпителий.
При увеличении количества желтка в яйце дробление Eutomostraca меняет свой характер. Так, в яйцах ракушковых (Ostracoda. рис. 135) и в зимних яйцах Daphina дробление остается полным до гаструляции, но после гаструляции (путем иммиграции) внутренние клетки сливаются, образуя желточную внутреннюю массу с ядрами. У Ostracoda ядра, в желтке, располагаются при этом на правильных расстояниях друг от друга, так что возможно, что полного исчезновения границ клеток и не происходит, но в зимних яйцах Daphina ядра во внутренней желточной массе рассеяны беспорядочно, чтоговорит о несомненном слиянии клеток. Клетки наружного слоя — эктодермы — не сливаются друг с другом, образуя наружный эпителий.
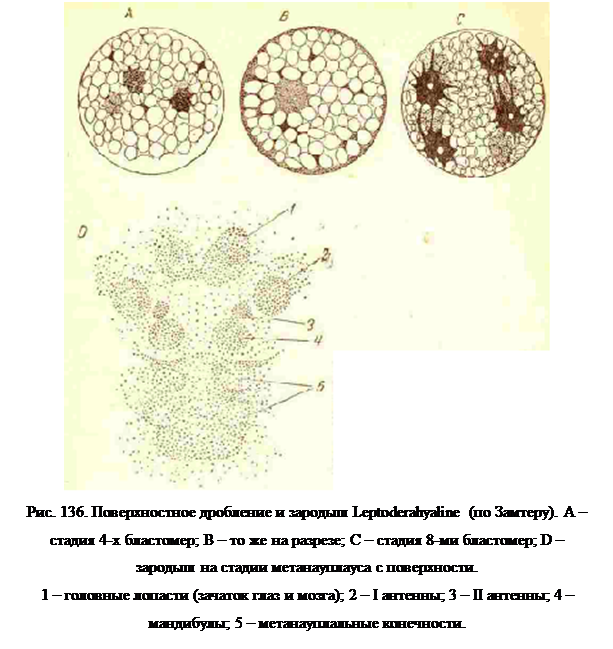
Среди Entomostraсa наибольшим количеством желтка обладают яйца Leptodora (из Ciadocera), в которых дробление с самого начала идет как поверхностное (рис. 136). Ядро, окруженное протоплазмой, лежит в центре яйца и протоплазматическими тяжами соединено с поверхностной бластемой; все промежутки между тяжам заполнены желтком. На стадии 2 и 4 бластомер ядра лежат внутри желтка, но к третьему делению они уже выходят на поверхность и начинают образовывать бластодерму. Клетки последней размножаются делением и мельчают, и ядра их в большой части стягиваются наодну сторону яйца, где образуют сгущение мелких ядер, из которого возникает зародыш.
 Из яиц с небольшим количеством желтка развивается характерная личинка науплиус. Она имеет мелкие, часто микроскопические размеры, тело ее короткое, яйцевидной формы и снабжено только тремя парами конечностей, составляющими только переднюю часть комплекта конечностей взрослого животного, т. е. антеннулы, антенны и мандибулы (ряс. 137. А). Из них только первая пара одноосная, которую можно сравнить с антеннулами взрослого, антенны же и мандибулы несут иную функцию, чем у взрослого, приспособлены для плавания и расщеплены в дистальной своей части на две ветви. Из теменной пластинки илиголовной лопасти зародыша образуется у науплиуса головной мозг и непарный глаз. Имеется пара нефридиев, по характеру своего канала очень сходных с протонофридиями аннелид; они открываются у основания второй пары конечностей. В этой же части тела находится начальная часть нефридия, так что последний образует петлю, которая своейвершиной доходит до заднего уровня третьей, последней пары науплиальных конечностей. Внутренняя метамерия в теле науплиуса не выражена, так как мезодерма частью уходит в зачатки конечностей, частью дает туловищную мускулатуру этих конечностей, располагающуюся продольно в виде длинных тяжей.
Из яиц с небольшим количеством желтка развивается характерная личинка науплиус. Она имеет мелкие, часто микроскопические размеры, тело ее короткое, яйцевидной формы и снабжено только тремя парами конечностей, составляющими только переднюю часть комплекта конечностей взрослого животного, т. е. антеннулы, антенны и мандибулы (ряс. 137. А). Из них только первая пара одноосная, которую можно сравнить с антеннулами взрослого, антенны же и мандибулы несут иную функцию, чем у взрослого, приспособлены для плавания и расщеплены в дистальной своей части на две ветви. Из теменной пластинки илиголовной лопасти зародыша образуется у науплиуса головной мозг и непарный глаз. Имеется пара нефридиев, по характеру своего канала очень сходных с протонофридиями аннелид; они открываются у основания второй пары конечностей. В этой же части тела находится начальная часть нефридия, так что последний образует петлю, которая своейвершиной доходит до заднего уровня третьей, последней пары науплиальных конечностей. Внутренняя метамерия в теле науплиуса не выражена, так как мезодерма частью уходит в зачатки конечностей, частью дает туловищную мускулатуру этих конечностей, располагающуюся продольно в виде длинных тяжей.
Дальнейшее развитие науплиуса состоит в росте всего тела, в особенности в длину при этом образуются в последовательности спереди назад новые пары конечностей, число которых различно у различных Entomostracаи которые называются метанауплиальными (рис. 137, В). Метанауплиальные сегменты с их конечностями возникают телобластически из эктодермальной зоны нарастания несколько впереди анального отверстиия Следовательно, источники возникновения науплиальной и метанауплиальной мезодермы так же различны, как и источники возникновения ларвальной и постларвальной мезодермы у аннелид. Кроме того, между этими обеими группами сегментов у Annelides и Entomostraca проявляется параллелизм в последовательности возникновения постларвальпых (метанауплиальных) сегментов и в единовременности появления ларвальных (науплиальных), а также в том, что сегментация мезодермы постларвального (метанауплиального) отдела тела выражена несравненно яснее, чем в науплиальных.
Рост науплиуса, сопровождаемый переходом его в следующую личиночную стадию — метапауплиуса, при наличии хитинового покрова, возможен только благодаря линьке, т. е. периодическому сбрасыванию хитина и образованию нового, и ограничен ею, так как видимое изменение и увеличение тела наступает только после линьки, когда сбрасывается стесняющий изменения тела хитиновый покров. Но на самом деле изменениеи рост идут непрерывно и постепенно, так что перед линькой ткани бывают несколько сжаты или смяты. Метанауплиус циклопа переходит в циклопидную стадию, похожую, но не вполне, на взрослого циклопа. Число линек в тех случаях или в те периоды, когда изменяется форма и длина тела и образуются новые конечности, совпадает с числом личиночных стадий; в те же периоды, когда личинкаравномерно растет, линька наступает, но не сопровождается переходом в новую личиночную стадию.
Наиболее сложной сменой личиночных стадий отличаются низшие ракообразные, ведущие сидячий или паразитический образ жизни.
Усидячих и паразитических усоногих науплиус превращается в метанауплиус, а последний — в так называемую циприсовидную личинку, снабженную двустворчатым панцирем или раковинкой, подобно Ostraсoda. У сидячих форм такая личинка прикрепляется к субстрату при помощи своих антеннул, снабженных на концах присоской и особой «цементной» железой. Сначала задний конец личинки направлен кверху, но затем, путем сгибания и образования складки в голове (рис. 138), животное поворачивается на 900, так что кверху теперь направлена брюшная сторона его. Участок головы впереди складки вырастает кверху ввиде толстого цилиндрического столбика (Lepas) или распластывается по субстрату (Balanus). У паразитического Sacculina циприсовидная личинка прикрепляется своими антеннулами к наружным  покровам молодых крабов в тех местах, где хитин толо- (суставы, основания волосков), после чего внутренние органы личинки подвергаются гистолизу и превращаются в клеточную массу; последняя стягивается к антеннулам, и наступает линька, причем лишенная содержимого задняя, часть тела отбрасывается. Оставшаяся у антеннул клеточная масса одевается затем хитином и принимает вид яйцевидного мешка, полость которого продолжается в полость антеннул, внедренных под покровы хозяина. По аптеннулам, имеющим вид хитиновых трубок, клеточная масса проникает в полость тела хозяина и ложится на вентральной стенке кишечника в его абдомене в виде скопления с корневидно расходящимися от нею клеточными тяжами, которые своими ветвлениями вплетают все органы тела. В центральном скоплении диференцируется мантия, надглоточный ганглий и огромный яичник, и все это скопление доходит до наружных покровов хозяина, прорывается наружу и высовывается и виде мешка, из которого выходят в воду яйца, превращающиеся в науплиусов.
покровам молодых крабов в тех местах, где хитин толо- (суставы, основания волосков), после чего внутренние органы личинки подвергаются гистолизу и превращаются в клеточную массу; последняя стягивается к антеннулам, и наступает линька, причем лишенная содержимого задняя, часть тела отбрасывается. Оставшаяся у антеннул клеточная масса одевается затем хитином и принимает вид яйцевидного мешка, полость которого продолжается в полость антеннул, внедренных под покровы хозяина. По аптеннулам, имеющим вид хитиновых трубок, клеточная масса проникает в полость тела хозяина и ложится на вентральной стенке кишечника в его абдомене в виде скопления с корневидно расходящимися от нею клеточными тяжами, которые своими ветвлениями вплетают все органы тела. В центральном скоплении диференцируется мантия, надглоточный ганглий и огромный яичник, и все это скопление доходит до наружных покровов хозяина, прорывается наружу и высовывается и виде мешка, из которого выходят в воду яйца, превращающиеся в науплиусов.
Если яйца содержат большое количество желтка, то стадия иауилпуса проходит в яйце, и вылущение происходит на стадии метанауплиуса. У Ostraсoda, яйца которых содержат довольно много желтка, из яйца хотя и выходит науплиус, но он отличается от обычной формы науплиуса образованием двустворчатого панцыря, делающего его похожим на взрослую форму Ostraсoda. Из богатых желтком зимних яиц Cladocera выходят уже вполне сформированные, по мелкие рачки. У Leptodora и дафниq и стадия науплиуса и стадия метанауплиуса проходит в яйце (рис. 136, D).
 Так как при большом количестве желтка дробление переходит к поверхностному типу, то и дальнейшее развитие, в частности образование конечностей, идет несколько иначе, чем в яйцах с малым количеством желтка. В последних вся масса состоит из живых и активных бластомер, и яйцо постепенно переформировывается целиком п тело паушшуса. При поверхностном же дроблении активные в отношении морфогенеза клетки находятся только на поверхности яйца, т. е. в бластодерме, а на ранних стадиях закладки органов ограничены даже только частью поверхности яйца, являющейся вентральной стороной его. Поэтому зачатки головных лопастей и конечностей появляются распластанными на поверхности желтка (рис. 136, D и 139, В), и только к концу эмбрионального развития, когда этот пассивный желток резорбируется при питании зародышевых частей, зародыш получает ту форму, которая определяется морфогенетической деятельностью его клеток.
Так как при большом количестве желтка дробление переходит к поверхностному типу, то и дальнейшее развитие, в частности образование конечностей, идет несколько иначе, чем в яйцах с малым количеством желтка. В последних вся масса состоит из живых и активных бластомер, и яйцо постепенно переформировывается целиком п тело паушшуса. При поверхностном же дроблении активные в отношении морфогенеза клетки находятся только на поверхности яйца, т. е. в бластодерме, а на ранних стадиях закладки органов ограничены даже только частью поверхности яйца, являющейся вентральной стороной его. Поэтому зачатки головных лопастей и конечностей появляются распластанными на поверхности желтка (рис. 136, D и 139, В), и только к концу эмбрионального развития, когда этот пассивный желток резорбируется при питании зародышевых частей, зародыш получает ту форму, которая определяется морфогенетической деятельностью его клеток.
Подобное расположение эмбриональных тканей встречается и у аннелид в том случае, если бластомеры вегетативного полушария сильно загружены желтком, как это имеет место, например, у Nereis, хотя у этой полихеты поверхностное размещение зачатков обусловлено не поверхностным дроблением, а, эпиболическим распространением микромер и их производных по поверхности желточных макромер основного квартета. Соответствие частей тела зародыша аннелиды и ракообразного с поверхностным дроблением выступает особенно отчетливо при сравнении их между собою (рис. 139). Перед зачаткомротового отверстия лежит теменная пластинка, которая у аннелид отделена от переднего края рта венцом крупных трохобластов; последние унекоторых Сopepoda заменяются кольцевидным органом, так называемым боковым или спинным органом, о котором будет сказано ниже. Позади ротового отверстия у Nereis лежат зачатки пары пальп и трех нар параподий ларвальных сегментов, а у Crustacea — трех пар науплиальных придатков, из которых первая пара (антеннулы) отличается от антенн аннелид присутствием в ней полости тела, а от параподий аннелид отсутствием двурасщепленности. Антеннулы, вероятно, являютсягомологами пальп, которые у аннелид закладываются по сторонам ротового отверстия, но позднее сдвигаются вперед и становятся придатками головы (т. е. prostoinium или предротовой лопасти). Внутреннее сходство выражается в том, что у Nereis макромеры сливаются в общую желточную массу, отделив от себя последнюю группу мелких клеток, из которых и образуется кишечник червя, а ядра макромер остаются в желтке и способствуют резорбции желтка у зародыша. У Crustacea с поверхностным дроблением образуется особый зачаток кишечника взрослого, который медленно развивается и диференцируется, а ядра, оставшиеся в желтке после образования бластодермы, служат для резорбции желтка и носят название вителлофагов.