 Первичнотрахейные стоят весьма близко к вымершей группе Protonychophora одной из самых древних, встречающейся в отложениях прекембрийской эпохи. Этой группой, представители которой совмещают признаки аннелид и членистоногих, начинается третий ствол Arthropoda, который может быть назван Tracheata. Среди Onychophora имеются виды, развивающиеся с большим количеством желтка, c малым количеством желтка и с плацентарным питанием зародыша. Из яиц c большим количеством желтка выходят маленькие особи (около 2 мv), а при плацентарном питании рождающееся животное достигает 20 мм.
Первичнотрахейные стоят весьма близко к вымершей группе Protonychophora одной из самых древних, встречающейся в отложениях прекембрийской эпохи. Этой группой, представители которой совмещают признаки аннелид и членистоногих, начинается третий ствол Arthropoda, который может быть назван Tracheata. Среди Onychophora имеются виды, развивающиеся с большим количеством желтка, c малым количеством желтка и с плацентарным питанием зародыша. Из яиц c большим количеством желтка выходят маленькие особи (около 2 мv), а при плацентарном питании рождающееся животное достигает 20 мм.
Развитие c большим количеством желтка (Peripatusnovac-zealandiac). Дробление поверхностное, но ядра из желтка выходят не на всю поверхность его, а только на одни полюс, где образуется пластинка бластодермы; отсюда происходит обрастание бластодермой всей поверхности яйца. Бластодерма на том полюсе, где она возникла, утолщается, а в утолщении появляется бластопор в виде прямой, продольной щели, дно которой занято желтком с рассеянными, оставшимися в нем ядрами (рис. 159). Дальнейшим утолщением краев щелевидного бластопора и пролиферацией из них клеток, образуются правая и левая зародышевые полоски (рис. 159, D); поверхностный слой полоски является эктодермой, прилегающий к желтку — энтодермой, мезодермальнымиже клетками будут все клетки, лежащие между эктодермой и энтодермой. По сторонамщелпи бластопора мезодерма образует мезодермальные полоски, которые сегментируются на пять пар сомитов, причем сегментация идет от заднего конца к переднему. Затем края бластопора срастаются, оставляя открытым лишь отверстие на переднем конце его, которое представляет собой ротовое отверстие; запаздывает и закрытие заднегоконца щели, и считается, что остающееся здесь отверстие является анальным. Позади него несколько ранее образуется утолщение, которое затем начинает пролиферировать мезодерму и энтодерму следующих сегментов, появляющихся значаительно позже и обособляющихсяв последовательности отпереднего конца к заднему.
 Развитие с малым количеством желтка (Peripatuscapensis). Желток находится в середине яйца, снаружи располагается хорошо выраженный слойпротоплазмы с ядром.Дробится только протоплазма, а бластомеры остаются соединенными с желтком.Бластомеры анимального полушария дают впоследствии эктодерму, а более крупныебластомеры вегетативного — энтодерму. Желток разжижается и яйцо (бластула)вытягивается по одной оси; затем бластула, вследствие потребления желтка, сильно съеживается, причем энтодерма сбивается в кучкуклеток, которую затемобрастает эктодерма. На стороне вегетативного полюса остается при этомузкая щель, представляющая собой бластопор (рис. 160). Образование из бластопора ротового и анального отверстий описаноисследователями сходно с этим процессов у P. novae-zealandiae Кроме переднего и заднего концов бластопора остается открытой и средняя часть на уровне 5-го сегментатела. Зачаток мезодермы возникает не по бокам бластопора, а позади него.
Развитие с малым количеством желтка (Peripatuscapensis). Желток находится в середине яйца, снаружи располагается хорошо выраженный слойпротоплазмы с ядром.Дробится только протоплазма, а бластомеры остаются соединенными с желтком.Бластомеры анимального полушария дают впоследствии эктодерму, а более крупныебластомеры вегетативного — энтодерму. Желток разжижается и яйцо (бластула)вытягивается по одной оси; затем бластула, вследствие потребления желтка, сильно съеживается, причем энтодерма сбивается в кучкуклеток, которую затемобрастает эктодерма. На стороне вегетативного полюса остается при этомузкая щель, представляющая собой бластопор (рис. 160). Образование из бластопора ротового и анального отверстий описаноисследователями сходно с этим процессов у P. novae-zealandiae Кроме переднего и заднего концов бластопора остается открытой и средняя часть на уровне 5-го сегментатела. Зачаток мезодермы возникает не по бокам бластопора, а позади него.
 Развитие плацентой (Peripatus edwardsii). Яйца мелкие; дробление полное и равномерное, и дает плотное скопление клеток, расположенных по радиальному типу, заполняющее просвет матки. Затем клетки выделяют из себя жидкость и сильно уменьшаются в размере, и все скопление их принимает теперь вид плотной однослойнойпластинки (рис. 161,В, 2), прижатой к стейке матки и срастающейся своими краями с ее эпителием; просвет матки снова делается при этом свободным.
Развитие плацентой (Peripatus edwardsii). Яйца мелкие; дробление полное и равномерное, и дает плотное скопление клеток, расположенных по радиальному типу, заполняющее просвет матки. Затем клетки выделяют из себя жидкость и сильно уменьшаются в размере, и все скопление их принимает теперь вид плотной однослойнойпластинки (рис. 161,В, 2), прижатой к стейке матки и срастающейся своими краями с ее эпителием; просвет матки снова делается при этом свободным.
. Пластинка растет в полость матки и изгибается в виде цилиндрического колпачка с узкой полостью внутри (рис. 161, С). Этот период соответствует периоду обрастания желтка бластодермойу P. novae-zealandiae. Прикрепленный к стенке, матки конец плода отвечает анимальному полюсу, а противоположный — вегетативному, но здесь вместо щелевидного бластопора появляется только неглубокая бороздка; то, что бороздка эта есть бластопор,доказывается усиленной миграцией из ее дна в полость плода клеток (рис. 162, В, 5),дающих энтодерму, которая принимает вид внутреннего эпителиального мешочка, имезодерму (рис. 162, В, 4). Плодрастет по одной оси, параллельной стенке матки, и соответственно удлиняется и энтодермальный и мезодермальный зачатки. Образование мезодермальных полосок идет так же, как и у других видов, итак же, как у них, появляютсясначала только 5 сомитов.Вследствие того что растеттолько дистальная часть плода,проксимальная его часть получает характер прикрепительной ножки, или пуповины(рис. 162), которая одним своим концом прикреплена к кольцевидному утолщению эпителияматки. Через это утолщение хорошо проникают в пуповинную ножку и в зародыша питательные вещества из кровиматери, и потому оно являетсяплацентой.
 От поверхности плода в начале роста зародыша выходит поодиночке клетки, образующие тонкий эпителиальный покров, который представляет собою амнион, но серозная оболочка, сопутствующая амниону у других животных, не появляется.
От поверхности плода в начале роста зародыша выходит поодиночке клетки, образующие тонкий эпителиальный покров, который представляет собою амнион, но серозная оболочка, сопутствующая амниону у других животных, не появляется.
В дальнейшем развитии всех Onychophora характерной особенностью являетсяболее длительное, чем у других Arthropoda, сохранение, целомическими метками целостности и замкнутости, что также сближает их с аннелидами. Готовые части лежат сначала на поверхности зародыша, как и остальные конечности, но позднее около нихобразуется складка, которая отчасти прикрывает их с боков.
В отношении описанных выше более ранних стадий развития Pеripatus необходимоуказать, что в них кое-что осталось невыясненным. Так, анальное отверстие считается задним остатком щелевидного бластопора, зона же нарастания новых сегментов телаописывается лежащей позади бластопора; неясно, каким образом в таком случае анальное отверстие оказывается затем в самом заднем из нарастающих сегментов. Крометого, одними авторами описано возникновение мезодермы и энтодермы из боковыхкраев бластопора (P. novae-zealandiae), другими — позади бластопора (у Р. capensis).
На основании сопоставления с развитием многоножек (Chilopoda) и с особенностямиметамерии аннелид и членистоногих можно предположительно, до дальнейших исследований развития Onyсhophora, представить этот процесс в следующем виде, отчастипримиряющем указанные противоречияи неясности. Из боковых краев щелевидногобластопора возникают энтодерма имезодерма головной лопасти и пятипередних сегментов, в остальныхлее сегментах образование мезодермы происходит деятельностью зоны нарастания, появляющейся несколько впереди заднего конца щели бластопора; этот конец егосмещается вследствие образованияновых сегментов но зоны роста, что сопровождается сильным удлинением бластопора. При замыкапни последнего остается открытым только его передний конец,т. е. ротовое отверстие; кроме того,иногда временно сохраняется щельна уровне 5-го сегмента (рис. 160, В)и остается или заново появляетсяотверстие в заднем конце бластопора, являющееся анальным илежащее в последнем сегменте.
Многоножки (Myriapoda)
Среди многоножек наиболеепримитивными в морфологическом отношении являются формы с малым числом; сегментов и еще меньшим числом пар ножек (не более 12).Из таких многоножек в особенностипредставители отряда Symphуla(сколопендреллы) обладают признаками, позволяющими считать ихблизкими к тем исходным формам, от которых произошли как остальные многоножки (Diplopoda, Cliilopoda), так и насекомые.
 Symphуla. У Hanseniella, австралийского представителя Sympliyla, желток равномерно распределен по всему яйцу; дробление яйца полное и почти равномерное(рис. 163, А), в результате чего получается бластула с хорошо выраженным бластоцелем. Последний со стадии 100—120 бластомер начинает частично заполняться бластомерами, так как с этой стадии некоторые деления в бластуле начинают происходить тангентально (рис. 163, В), В бластоцеле эти внутренние бластомеры затем утрачивают свои границы и сливаются во внутреннюю сплошную массу желтка, а ядра егос небольшими участками протоплазмы становятся желточными клетками. Наружные же клетки бластулы сохраняют характер эпителия, беднеют желтком и становятсяклетками бластодермы; они имеют столбчатую форму, ядра же их при дальнейших делениях постепенно смещаются к самой поверхности яйца.
Symphуla. У Hanseniella, австралийского представителя Sympliyla, желток равномерно распределен по всему яйцу; дробление яйца полное и почти равномерное(рис. 163, А), в результате чего получается бластула с хорошо выраженным бластоцелем. Последний со стадии 100—120 бластомер начинает частично заполняться бластомерами, так как с этой стадии некоторые деления в бластуле начинают происходить тангентально (рис. 163, В), В бластоцеле эти внутренние бластомеры затем утрачивают свои границы и сливаются во внутреннюю сплошную массу желтка, а ядра егос небольшими участками протоплазмы становятся желточными клетками. Наружные же клетки бластулы сохраняют характер эпителия, беднеют желтком и становятсяклетками бластодермы; они имеют столбчатую форму, ядра же их при дальнейших делениях постепенно смещаются к самой поверхности яйца.
 Затем в бластодерме появляется широкая зародышевая полоска, занимающая большую часть поверхности яйца. Мезодерма возникает выклиниванием клеток из всех частей зародышевой полоски и слагается в две мезодермальные полоски. Вскоре после появления зародышевой полоски начинается бластокинез, выражающийся в образовании глубокой поперечной борозды или перегиба по середине полоски (рис. 164, А). В то же время идет обособление первых сегментов; первым появляется сегмент нижней губы на самой глубокой части перегиба (рис. 164, В, 5). Затем следует образованиевпереди губного сегмента максиллярного, мандибулярного, премандибулярного (или интеркалярного) сегментов, а плзади губного обособляются сегменты туловища. Всего образуется девять туловищных сегментов, остальные семь возникают уже в постэм брионалышй период.
Затем в бластодерме появляется широкая зародышевая полоска, занимающая большую часть поверхности яйца. Мезодерма возникает выклиниванием клеток из всех частей зародышевой полоски и слагается в две мезодермальные полоски. Вскоре после появления зародышевой полоски начинается бластокинез, выражающийся в образовании глубокой поперечной борозды или перегиба по середине полоски (рис. 164, А). В то же время идет обособление первых сегментов; первым появляется сегмент нижней губы на самой глубокой части перегиба (рис. 164, В, 5). Затем следует образованиевпереди губного сегмента максиллярного, мандибулярного, премандибулярного (или интеркалярного) сегментов, а плзади губного обособляются сегменты туловища. Всего образуется девять туловищных сегментов, остальные семь возникают уже в постэм брионалышй период.
У Hanseniellaимеется на спинной стороне бластодермы дорзальный орган; в отличие от дорзальных органов других членистоногих он выполняет свою функцию отслаивания хитина при эмбриональных линьках выделением не гигроскопического, коллоидального вещества, а плотных нитей (рис. 163, С), которые достигают значительной длины и образуют целое сплетение между эмбриональной кутикулой и эктодермой.

 Двупарноногие (Diplopoda). У некоторых представителей (Polydеsmus) дробление вначале полное (рис. 165), по на шестом делении протоплазма стягивается на поверхность, и границы поверхностных бластомер исчезают, равно как и границы внутренних бластомер, протоплазма которых стягивается к их ядрам; дробление переходит тогда в поверхностное (рис. 166). У других представителей (Plathyrraсus) дробление с самого начала поверхностное. В желтке остаются некоторые ядра и способствуют его усвоению, образуя желточную энтодерму. Кишечная энтодерма появляется из того участка бластодермы, из которого несколько позднее впячивается эктодермальный стомодеум; клетки этой энтодермы не теряют связи друг с другом и сначала в виде рыхлого, затем плотного тяжа внедряются в желток и растут и нем параллельно вентральной стороне яйца к заднему концу зародышевого пятна (рис. 166, В). Они не захватывают в себя желтка, который до конца развития остается между эктодермой и энтодермой, т. е. заполняет первичную полость тела. Мезодерма возникает из двух симметричных участков бластодермы, из которых выходят под бластодерму клетки (рис. 166,2); последние распространяются под эктодермальным зародышевым пятном и вскоре разделяются на две мезодермальиые полоски. Затем начинается сегментация эктодермы в виде местных парных сгущений ее клеток.
Двупарноногие (Diplopoda). У некоторых представителей (Polydеsmus) дробление вначале полное (рис. 165), по на шестом делении протоплазма стягивается на поверхность, и границы поверхностных бластомер исчезают, равно как и границы внутренних бластомер, протоплазма которых стягивается к их ядрам; дробление переходит тогда в поверхностное (рис. 166). У других представителей (Plathyrraсus) дробление с самого начала поверхностное. В желтке остаются некоторые ядра и способствуют его усвоению, образуя желточную энтодерму. Кишечная энтодерма появляется из того участка бластодермы, из которого несколько позднее впячивается эктодермальный стомодеум; клетки этой энтодермы не теряют связи друг с другом и сначала в виде рыхлого, затем плотного тяжа внедряются в желток и растут и нем параллельно вентральной стороне яйца к заднему концу зародышевого пятна (рис. 166, В). Они не захватывают в себя желтка, который до конца развития остается между эктодермой и энтодермой, т. е. заполняет первичную полость тела. Мезодерма возникает из двух симметричных участков бластодермы, из которых выходят под бластодерму клетки (рис. 166,2); последние распространяются под эктодермальным зародышевым пятном и вскоре разделяются на две мезодермальиые полоски. Затем начинается сегментация эктодермы в виде местных парных сгущений ее клеток.
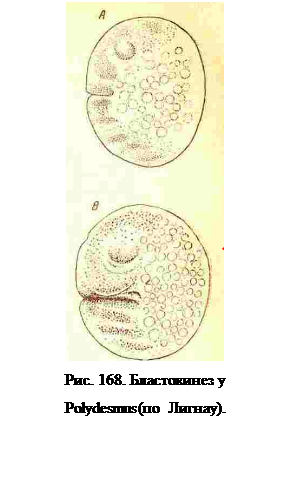
 Сгущение, появляющееся впереди стомодеума, дает головные лопасти; пара сгущений по сторонам стомодеума дает сегмент антенн. Позади антенн возникают еще мять ясно обособленных сегментов (рис. 167): мандибул, максилл, рудиментарный постмаксилл, первой и второй пар конечностей и расплывчатое пятно, которое позднее отделяют от себя вперед сегменты третьей и четвертой пар конечностей. На уровне сегментамаксилл затем возникает на вентральной стороне поперечная, входящая в желтокскладка, которая превращается в очень глубокую борозду, разделяющую яйцо на двеполовины. В эту складку втягиваются образовавшиеся сегменты, причем все головныесегменты (до первой лары ходных конечностей) входят в переднюю поверхность складки,а остальные в заднюю (рис. 168). Образованием складки выражен у Diplopoda процессбластокинеза. После потребления всего желтка из яйца вылупляется личинка, несущая только головные конечности и три, реже четыре пары ходных ножек (рис. 169).
Сгущение, появляющееся впереди стомодеума, дает головные лопасти; пара сгущений по сторонам стомодеума дает сегмент антенн. Позади антенн возникают еще мять ясно обособленных сегментов (рис. 167): мандибул, максилл, рудиментарный постмаксилл, первой и второй пар конечностей и расплывчатое пятно, которое позднее отделяют от себя вперед сегменты третьей и четвертой пар конечностей. На уровне сегментамаксилл затем возникает на вентральной стороне поперечная, входящая в желтокскладка, которая превращается в очень глубокую борозду, разделяющую яйцо на двеполовины. В эту складку втягиваются образовавшиеся сегменты, причем все головныесегменты (до первой лары ходных конечностей) входят в переднюю поверхность складки,а остальные в заднюю (рис. 168). Образованием складки выражен у Diplopoda процессбластокинеза. После потребления всего желтка из яйца вылупляется личинка, несущая только головные конечности и три, реже четыре пары ходных ножек (рис. 169).

 Превращение во взрослое животное происходит путем образования новых сегментов из зоны нарастания в задней части тела личинки, причем зона эта находится в эктодерме. Процесс этого анаморфоза сопровождается другими своеобразными процессами —образованием двойных сегментов. Онсостоит в том, что закладываетсяодин сегмент с одной парой ганглиев,но при первом появлении в нем зачатков конечностей каждый ганглий разделяется на два; и в таком сегменте соответственно закладываются две пары конечностей. Спереди и сзадиэтих двух пар вырастают большие складки эктодермы, которые срастаются друг с другом, образуя замкнутый карман для этих двух пар зачатков ног; при дальнейшем развитии сегмента складки атрофируются, а ноги вырастают. Такие двойные сегменты возникают на всем протяжении отдела тела, образующегося при постэмбриональном развитии, а частично двойными становятся и некоторые сегменты, возникшие во время эмбрионального развития.
Превращение во взрослое животное происходит путем образования новых сегментов из зоны нарастания в задней части тела личинки, причем зона эта находится в эктодерме. Процесс этого анаморфоза сопровождается другими своеобразными процессами —образованием двойных сегментов. Онсостоит в том, что закладываетсяодин сегмент с одной парой ганглиев,но при первом появлении в нем зачатков конечностей каждый ганглий разделяется на два; и в таком сегменте соответственно закладываются две пары конечностей. Спереди и сзадиэтих двух пар вырастают большие складки эктодермы, которые срастаются друг с другом, образуя замкнутый карман для этих двух пар зачатков ног; при дальнейшем развитии сегмента складки атрофируются, а ноги вырастают. Такие двойные сегменты возникают на всем протяжении отдела тела, образующегося при постэмбриональном развитии, а частично двойными становятся и некоторые сегменты, возникшие во время эмбрионального развития.
Губоногие (Chilopoda). Дробление яйца Geopholusи Seolopendra поверхностное, но вначале оно сопровождается образованием желточных пирамид, сливающихся между собой в центре яйца и разделенных тонкими прослойками протоплазмы (рис. 170). Затем границы желточных пирамид исчезают, и все ядра выходят на поверхность желтка, по некоторые из них остаются тесно связанными с желтком и являются желточной энтодермой, в которой лежат ядра дробления. Большая часть ядер образует бластодерму только на одной стороне яйца, и в центре ее в виде сгущения клеток возникает дисковидное зародышевое пятно (рис. 172, А). К одному краю пятна прилегает половой зачаток в виде хорошо очерченного сферического образования; по положению своему он занимает задний край пятна.
 В центре зародышевого пятна появляется слабо выраженная продольная первичная полоска, концы которой не доходят до краев диска. В этой полоске происходит миграция клеток из бластодермы к желтку по очень узкой срединной линии, где вследствиеэтого появляется очень неглубокая продольная бороздка (рис. 171, А). Часть выклинивающихся из нее клеток входит и слой желточной энтодермы, и, постепенно увеличиваясь с размерах, эти клеткистановятся сходными с клетками этой последней, большая же часть клеток бороздки расходится в стороны и образует мезодерму.
В центре зародышевого пятна появляется слабо выраженная продольная первичная полоска, концы которой не доходят до краев диска. В этой полоске происходит миграция клеток из бластодермы к желтку по очень узкой срединной линии, где вследствиеэтого появляется очень неглубокая продольная бороздка (рис. 171, А). Часть выклинивающихся из нее клеток входит и слой желточной энтодермы, и, постепенно увеличиваясь с размерах, эти клеткистановятся сходными с клетками этой последней, большая же часть клеток бороздки расходится в стороны и образует мезодерму.
Зародышевое пятно удлиняется, отодвигая половойзачаток все дальше назад, и вскоре пролиферационная деятельность срединной полоски прекращается, а засчет возникшей из нее мезодермы появляются шестьпередних сегментов (рис. 172, В и С):1) антеннальный,2) исчезающий  затем интеркалярный, 3) мандибулярный,4) первый максиллярный, 5) второй максиллярныйи 6) ногочелюстной. Этот комплект сегментов затемдолго не пополняется новыми, но развитие этих сегментов продолжается до появления на них первых зачатков конечностей. Появляется также и стомодеум. За
затем интеркалярный, 3) мандибулярный,4) первый максиллярный, 5) второй максиллярныйи 6) ногочелюстной. Этот комплект сегментов затемдолго не пополняется новыми, но развитие этих сегментов продолжается до появления на них первых зачатков конечностей. Появляется также и стомодеум. За 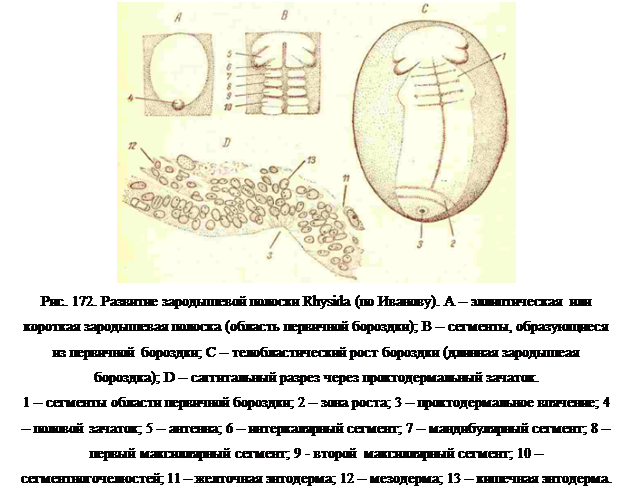 это времязародышевая полоска сильно удлиняется, но уже за счет другого источника, которыйимеет с поверхности вид поперечного утолщения недалеко от заднего конца зародышевой полоски и отодвигается далеко назад по мерероста полоски, сохраняя,однако, неизменное расстояние между собой изадним концом полоски споловым зачатком (рис.172, С, 2). Рост полоски в длину идет очень быстро, и вскоре появляются ввиде поперечных бороздок границы новых возникающих сегментов в правильной последовательностиспереди назад, причем самые передние из них резкоотличаются по степенисвоего развития от всех шести ранее образовавшихся сегментов. Помимо того что первичная полоска активна по всей своейдлине, а зона нарастания новых сегментов в продольном направлении активна в небольшом участке в заднем конце, характер их деятельности разнится, во-нервых, тем, что в зоне нарастания выклиниваине клеток происходит широким фронтом (рис. 171, В), а не только по срединной линии, и, во-вторых, тем, что и зоне нарастания образуются только клетки мезодермы. Из этой зоны возникают все сегменты, несущие ходные ложки, и зародышевая полоска настолько удлиняется, что передний и задний концы их очень близкосходятся друг с другом на дорзальной стороне яйца.
это времязародышевая полоска сильно удлиняется, но уже за счет другого источника, которыйимеет с поверхности вид поперечного утолщения недалеко от заднего конца зародышевой полоски и отодвигается далеко назад по мерероста полоски, сохраняя,однако, неизменное расстояние между собой изадним концом полоски споловым зачатком (рис.172, С, 2). Рост полоски в длину идет очень быстро, и вскоре появляются ввиде поперечных бороздок границы новых возникающих сегментов в правильной последовательностиспереди назад, причем самые передние из них резкоотличаются по степенисвоего развития от всех шести ранее образовавшихся сегментов. Помимо того что первичная полоска активна по всей своейдлине, а зона нарастания новых сегментов в продольном направлении активна в небольшом участке в заднем конце, характер их деятельности разнится, во-нервых, тем, что в зоне нарастания выклиниваине клеток происходит широким фронтом (рис. 171, В), а не только по срединной линии, и, во-вторых, тем, что и зоне нарастания образуются только клетки мезодермы. Из этой зоны возникают все сегменты, несущие ходные ложки, и зародышевая полоска настолько удлиняется, что передний и задний концы их очень близкосходятся друг с другом на дорзальной стороне яйца.
 Еще на довольно ранних стадиях деятельности зоны роста, то раньше, то позжена некотором расстоянии позади зоны появляется впячение, из которого впоследствиивозникает эктодермальныйпроктодеум (рис. 172, С и D, 3). В начале своего появления это впячение также проявляет усиленную пролиферационную деятельность, со дна его выклинивается довольно значительное количество клеток мезодермы, образующихпродолжение мезодермальных полосок, и не менее значительное количество клеток,которые ложатся на поверхность желточной эантодермы в виде плотной эпиталиальной пластинки (рис. 173, 3), последняя затем растет вперед и образует кишечную энтодерму сколопендры. Область проктодеума несколько приподимается над поверхностьюяйца, и у основания этого выступа образуются из возникшей здесь мезодермы еще тризадних сегмента, на которых появляются зачатки конечностей, но затем эти зачаткиисчезают, и три последних сегмента лишены ног.
Еще на довольно ранних стадиях деятельности зоны роста, то раньше, то позжена некотором расстоянии позади зоны появляется впячение, из которого впоследствиивозникает эктодермальныйпроктодеум (рис. 172, С и D, 3). В начале своего появления это впячение также проявляет усиленную пролиферационную деятельность, со дна его выклинивается довольно значительное количество клеток мезодермы, образующихпродолжение мезодермальных полосок, и не менее значительное количество клеток,которые ложатся на поверхность желточной эантодермы в виде плотной эпиталиальной пластинки (рис. 173, 3), последняя затем растет вперед и образует кишечную энтодерму сколопендры. Область проктодеума несколько приподимается над поверхностьюяйца, и у основания этого выступа образуются из возникшей здесь мезодермы еще тризадних сегмента, на которых появляются зачатки конечностей, но затем эти зачаткиисчезают, и три последних сегмента лишены ног.
Первичную полоску можно гомологизировать со щелевидным бластопором Оnуchophora как место пролиферации энтодермы и мезодермы и образования передних пяти пар конечностей и антенн, причем у Оnуchophora развиваются преантенны на дорзальной стороне головы, а у сколопендры развиваются антенны, расположенные в одном ряду о остальнымиконечностями. Затем ненаконце первичной полоски, а несколько впереди ее заднего конца возникает зона роста, дающая большую часть сегментов. Но еще до окончания деятельности этой зоны задний участок первичной полоски возобновляет свою активность в области проктодеума и дает еще три сегмента. Можно считать, что первичное тело Chilopoda (соответствующее ларвальному телу аннелид)состоит из восьми сегментов, не считая антенн.
Правая и левая зародышевые полоски при закладке на них конечностей отходят друг от друга, оставаясь соединенными только на переднем и заднем концах, и изгибаются крутой дугой в дорзальном направлении. Так начинается у Chilopoda бластокинез, который сопровождается образованием поперечной складки посередине зародышевых полосок, разделяющей желток на дне половины, причем передний и заднийконцы полосок смещаются к двум противоположным полюсам яйца; из первичной полоскиформируется изогнутой дугой зародыш более поздних стадий. У переднего конца зародышевых полосок описан на более ранних стадиях дорзальный орган в виде скопленияклеток внезародышевой эктодермы; существование его находится в связи с эмбриональными линьками, свойственными Chilopoda.
Описанный способ закладки зародышевых пластов свойствен некоторым представителям Chilopoda и является характерным. У других представителей, в частности, у родаScolopendra, описан несколько иной способ закладки пластов и сегментации зародышевой полоски, сходной отчасти с соответственным процессом Symphyla, отчасти с таковым некоторых насекомых.
 . Можно сказать, что закладка пластов и у многоножек и у насекомых значительно варьирует, но в обоих классах она варьирует сходно.
. Можно сказать, что закладка пластов и у многоножек и у насекомых значительно варьирует, но в обоих классах она варьирует сходно.