Основным отличием размножения Anamnia от Amniota служит то, что яйца Anamnia развиваются в воде, даже яйца тех из них, которые постоянно живут на суше (жабы, лягушки, саламандры). У Amniota же мы находим обратное: яйца всегда развиваются на суше, даже яйца тех из них, которые постоянно лижут в воде (черепахи и крокодилы). Это указывает па то, что в строении и развитии яйца Amniota выработались прочно установившиеся приспособления к развитию на суше.
К таким приспособлениям прежде всего относится прочная наружная скорлупа яйца, вырабатываемая стенками яйцеводов матери. Она имеет или кожистую консистенцию, или пропитана известью. Благодаря этому она достаточно прочна для того, чтобы,с одной стороны, предохранить живую часть яйца от механических поврежденийизвне, с другой — чтобы сдерживать жидкое или полужидкое содержимое яйца скорлупой, как стенками сосуда, ввиду большой разницы между удельным весом этого содержимого и воздуха. Кроме скорлупы могут быть еще дополнительные перепончатые илижидкие третичные оболочки; последние —в виде белка, который имеет значение дополнительного питательного материала. Помимо этих третичных оболочек яйца Amniotaснабжены или очень тонкой желточной или же вторичной оболочкой, вырабатываемой фолликулом (оолемма).
Белок, окружающий яйцо в скорлупе, не является, однако, благоприятной средойдля развития зародыша. Кроме того, содержание воды в белке может колебаться в зависимости от степени влажности воздуха. Еще менее благоприятно для развития зародышапрямое соприкосновение с оболочкой яйца, таккак на суше это может сопровождаться подсыханием оболочек от недостатка влажности и повреждением соприкасающегося с ними зародыша.Поэтому вскоре после начала развития около зародыша начинает развиваться амнион, т. е. зародышевая оболочка, которая покрывает его и наполнена жидкостью. Так же как у насекомых,вместе с амнионом возникает в виде наружной стороны складки, образующей амнион, серозная оболочка, составляющая одно целое с внезародышевой эктодермой.
Скорлупа яйца, пронизана сквозными порами,допускающими приток к яйцу кислорода и газообмен в зародыше. В связи с этим возникает вторая зародышевая оболочка, которая появляетсяв виде выроста вентральной стенки задней частикишечника и, быстро разрастаясь, прикрываетсобою не только зародыша, но и всю поверхностьжелтка. Так как этот эктодермальный вырост одетснаружи слоем мезенхимы, то в стенках его возникает богатая сеть кровеносных сосудов. Этасосудистая зародышевая оболочка, называемая аллантойсом (рис. 325), разрастается под серознойоболочкой и плотно к ней прижата, так что сосуды аллантойса ветвятся под самой поверхностьювсего яйца. Благодаря этому в них может легко идти газообмен, а густота сети сосудов и ее большая поверхность соприкосновения с серозной оболочкой не только околосамого зародыша, но и по всему яйцу обеспечивает достаточную интенсивность дыхания.
В связи с появлением амниона возникает еще новое образование, характерное для Amniota. Зародыш, одетый амнионом, лежит между серозной оболочкой и желтком, и амнион несколько вдавливается в желток. Поэтому по сторонам амниона получается промежуточное пространство между желтком и серозной оболочкой. Серозная оболочка изнутри, амнион и желток снаружи одеты слоем мезенхимы. Следовательно, эта промежуточная полость ограничена сверху мезенхимой серозной оболочки, снизу — мезенхимой желтка, а с третьей стороны — мезенхимой амниона. Так как полость, ограниченная со всех сторон мезодермой, называется целомом и в данном случае этот целом лежит вне тела зародыша, то он получил название экзоцелома (рис. 325, 10). Внутри экзоцелома и происходит разрастание аллантойса.
Помимо этих приспособительных особенностей яйца Amniota, в развитии зародыша также имеются некоторые общие для всех них отличия от Anamnia. Укажем пока только главнейшие из них.
У Anamnia хорда хотя и является у большинства из них провизорным органом, но, у всех форм, имеющих личинку, хорда выполняет у нее функцию опорного органа, так как дефинитивный скелет у личинки или совсем отсутствует, или развит очень слабо. Поэтому типичным для зародыша Anamnia признаком служит довольно значительнаятолщина хордального тяжа; таким он бывает даже у тех из них, которые имеют прямоеразвитие (селяхии, отчасти костистые рыбы). У Amniota же хорда никогда не функционирует как опорное образование у свободно двигающегося зародыша; у них это исключительно провизорный орган, появляющийся как основной зачаток, необходимый для возникновения всей организации. Поэтому у них хорда представляет собою тонкий и слабый тяж.
Другой особенностью развития зародыша всех Amniota является способ диференцировки миотомов в связи с обилием клеток в их мезенхиматозных производных.У Anamnia обращенная к хорде стенка миотома дает мускульную пластинку, обращенная к эктодерме дает мезенхиму кожи и только нижний внутренний угол миотома продуцирует склеротомную мезенхиму. У Amniota же после обособления миотома от спланхнотома начинает распадаться на мезенхимные клеткхг прежде всего та стенка миотома,где было до сих пор соединение со спланхнотомом (нижне-боковая), причем эта мезенхима входит в полость миотома и заполняет ее. Из этих клеток образуется только меньшая часть склеротомной мезенхимы, главная же ее масса получается распадением наклетки всей обращенной к хорде стенки миотома.Характер цельной пластинки сохраняет тольконаружная стенка миотома, обращенная к эктодерме, т. е. дерматом; ее верхний край являетсязачатком мускульной пластинки, которая получается разрастанием клеток этого края вниз повнутренней поверхности дерматома  (рис. 326, 4).Следовательно,возникает двойная кожно-мускульная пластинка, которая, разрастаясь в вентральном направлении, дает сплошной мускульный икожный слои всего тела; только во время этогоразрастания дерматом начинает распадаться наклетки кожной мезенхимы. Таким образом, междухордой и нервной системой,с одной стороны, имускульной пластинкой миотома, с другой,лежит мощная масса склеротомной мезенхимы(рис. 326,5), дающей дефинитивный позвоночник,а мускульная пластинка отодвинута отой мезенхимой к самой коже.
(рис. 326, 4).Следовательно,возникает двойная кожно-мускульная пластинка, которая, разрастаясь в вентральном направлении, дает сплошной мускульный икожный слои всего тела; только во время этогоразрастания дерматом начинает распадаться наклетки кожной мезенхимы. Таким образом, междухордой и нервной системой,с одной стороны, имускульной пластинкой миотома, с другой,лежит мощная масса склеротомной мезенхимы(рис. 326,5), дающей дефинитивный позвоночник,а мускульная пластинка отодвинута отой мезенхимой к самой коже.
Большое количество мезенхимы дает и сплапхнонлевра спланхнотомов; складки последней ипромежутки между нею и кишечником наполняются мезенхимными клетками. Частичному распадению на мезенхимоподобные клетки подвергаютсяи те мускульные пластинки, которые затем мигрируют в зачаток конечности и даютздесь ее мускулатуру. Образования мускульных почек у Amniota не происходит, чтообъясняется тем, что мускульные почки, достаточные для снабжения мускулатуройкоротких плавников и конечностей Anamnia, слишком малы и слабы, чтобы дать болееобъемистую и сложную мускулатуру более длинных и лучше развитых конечностейAmniota. Поэтому на образование ее у последних идут целиком мускульные пластинкиближайших к зачатку конечности миотомов, чем нарушается та однообразная метамерия мышечного аппарата, которая свойственна Anamnia.
В образовании выделительных органов у Amniota характерны слабое развитие иочень короткое существование предпочки, а также то, что при образовании первичнойпочки задние нефротомы ее области ие дают от себя нефридиальных канальцев, и образуют сплошную нефрогенную ткань, из которой затем формируется без метамернойпоследовательности большое количество нефридиальных канальцев. Самая же задняячасть этой нефрогенной ткани соединяется образующимися в ней нефридиальными канальцами не с первичнопочечным протоком, а с особыми мочеточниками, вырастающимииз заднего конца этого протока вперед и расширяющимися иа конце в широкие почечные лоханки, которые и принимают в себя выводные отверстия нефридиальных канальцев. Эти почки известны под названием вторичных почек.
Наконец, как на своеобразную особенность развития Amniota можно указать на парную закладку сердца. Но этот признак является лишь следствием очень рапней закладки первого зачатка сердца, которая наступает тогда, когда на уровне сердца кишечник еще не отделился от внезародышевой энтодермы и соединен с ней вертикальными связующими стенками; по сторонам этих стенок и появляются правый и левый зачатки эндокардия, которые сливаются в непарный эндокардий под кишечником после обособления последнего на уровне сердца от желточного мешка.
Из принадлежащих к Amniota рептилий, птиц и млекопитающих последние по условиям своего развития стоят особняком, так как развитие их происходит внутри тела матери с образованием плаценты. Поэтому рептилии и птицы в своем развитии имеют больше общего друг с другом, чем с млекопитающими, а так как у них также больше и морфологических черт сходства, то их объединяют в группу Sanropsida.
Однако, в ходе развития всех Amniota гораздо больше единообразия, чем в развитии различных представителей Anamnia, и только что сформированные зародыши всех представителей Amniota настолько похожи друг на друга, что, имея перед собой только тела зародыша без его зародышевых оболочек, очень трудно бывает сказать, к какому классу он принадлежит. Первые стадии закладки зародыша представлены двумя основными вариантами, одни из которых свойственен птицам, другой рептилиям. Однако, и в этих вариантах нет принципиальных различий, и у млекопитающих мы встречаем оба варианта. Стадии дробления уже существенно отличаются у Sanropsida от млекопитающих, но эти стадии вообще находятся иод наибольшим влиянием условий развития и количества желтка. Однако, хотя у млекопитающих дробление полное, а у Sauropsida дискои- дальное, характер развития остается сходным и полное дробление млекопитающих гораздо легче сравнивать с дроблением Sanropsida, чем с полным же дроблением, например ланцетника.
Для большинства Anamnia основным типом их развития, притом наиболее изученным, могут служить амфибии, и описание развития Anamnia Gnathostomata мы начали с амфибий, хотя филогенетически они и не являются примитивной группой. Таким же общим исходным типом, который проявляется еще более единообразно у всех Amniota н изучение которого уже насчитывает несколько столетий, является развитие птиц, в частности куриного зародыша, и описание развития Amniota мы начнем с этих представителей Sanropsida, хотя наиболее примитивными из них являются рептилии.
ГЛАВА XXXII. SAUROPSIDA
Яйцо очень богато желтком и имеет вид большого желточного шара с маленьким зародышевым пятном на анималыюм полюсе. Желток имеет неоднородный характер и состоит из чередующихся концентрических слоев более светлого и более темного желтка; светлый желток образует кроме того центральное скопление (пандерово ядро), от которого идет тяж к зародышевому пятну. Анпмальный полюс с зародышевым пятном удельно легче остального яйца, так что при изменении положения яйца он всегда поворачивается к верху.
Готовое яйцо выходит из яичника в яйцевод через широкую воронку последнего, одетое только тонкой желточной оболочкой. В яйцеводе яйцо оплодотворяется. Проходя по яйцеводу дальше, оно одевается еще третичными оболочками: тонкой волокнистой оболочкой, прикрепляющейся к желточной, а также белком или белковой оболочкой различной толщины. В конце яйцевода особые железы выделяют довольно прочную наружную пергаментную оболочку и скорлупу, у одних представителей содержащую известь, у других лишенную ее и состоящую из прочного волокнистого вещества.
Вскоре после оплодотворения начинается дробление в бластодиске, и в зависимости от того, насколько быстро у того или другого вида продвигается яйцо по яйцеводу, оно может откладываться или на стадиях самого начала дробления или на любой из последующих стадий развития вплоть до самых последних. Особенно большие видовые вариации имеют место у различных представителей рептилий; у птиц же яйцо откладывается или на стадиях начала дробления (голубь) или конца дробления (курица).
Птицы (Aves)
 Яйцо и дробление. Яйцо поддерживается в белке на неизменном расстоянии отскорлупы при помощи двух отходящих от противоположных его полюсов закрученныхтяжей желатинизироваииого белка, допускающих вращение яйца в белке (халазы).Яйцо вращается только вокруг длинной оси скорлупы; поэтому можно констатироватьчто направление осей самого зародышас самого начала детерминировано по отношению к осям скорлупы, именно так, чтопри лежачем положении яйца, когда тупой конец скорлупы обращен налево,а более острый направо, зародыш направлен передним концом от наблюдателя,а задним к нему (рис. 347).
Яйцо и дробление. Яйцо поддерживается в белке на неизменном расстоянии отскорлупы при помощи двух отходящих от противоположных его полюсов закрученныхтяжей желатинизироваииого белка, допускающих вращение яйца в белке (халазы).Яйцо вращается только вокруг длинной оси скорлупы; поэтому можно констатироватьчто направление осей самого зародышас самого начала детерминировано по отношению к осям скорлупы, именно так, чтопри лежачем положении яйца, когда тупой конец скорлупы обращен налево,а более острый направо, зародыш направлен передним концом от наблюдателя,а задним к нему (рис. 347).
В яйцах так называемых птепцовыхптиц белок по своему составу и свойствам отличается от белка выводковых и содержит меньше питательных веществ. Поэтому у птенцовых из яйца вылупляютсяптенцы, лишенные перьев и не способныеходить и самостоятельно клевать корм,к чему вполне готовы вылупляющиесявыводковые птицы. Вследствие недоразвитости птенцовых птиц при вылуплениии необходимости жить в таком состояниивне яйца у них имеются в этот периодспециальные временные приспособления,придающие им личиночный характер,как, например, светящиеся пятна покраям рта у некоторых птенцов, живущих в темных дуплах деревьев.
В яичнике (например, курицы) находятся ооциты различной величины и возраста, одетые фолликулом; в периоджелткообразования фолликул обильнооплетен сетью кровеносных сосудов; наполюсе фолликула, противоположномполюсу его прикрепления к яичнику,остается полоска, лишенная сосудов иназываемая цикатрикулой (рис. 327, 11).Когда ооцит достигнет окончательныхразмеров, цикатрикула фолликула разрывается, и яйцо попадает через полостьтела в воронку яйцевода (10). В начальной части последнего скопляются сперматозоиды после спаривания. В яйцопроникает обычно несколько сперматозоидов, но оплодотворяет его только один.
У курицы через 2—3 часа после оплодотворения яйцо попадает в железистый отдел яйцевода, где выделяется его стенками плотный белок и халазы, а в бластодиске происходит первое деление дробления. Через 3—4 часа после оплодотворения яйцо приходит в нижний конец яйцевода, где выделяется жидкий белок и скорлуповая перепонка, бластодиск же делится на 8 бластомер. Далее яйцо проходит в так называемую матку (самая нижняя часть яйцевода), в которой продолжается выделение жидкого белка ивыделяется скорлупа; здесь яйцо задерживается от 4-го до 21-го или даже до 27-го часа и откладывается на стадии начала образования энтодермы, а при большей задержке в матке — к концу этого процесса.
Дробление дискоидальное, и до четвертого или пятого делений борозды проходят перпендикулярно к поверхности. Некоторое отставание в делении периферическойчасти бластодиска наблюдаемое у Anamnia, у Amniota выражено чрезвычайно сильно,,
 так что в течение большей части дробления делится только центральная область бласто-
так что в течение большей части дробления делится только центральная область бласто-
диска (рис. 328). Направлениеборозд только при втором делении перпендикулярно к предыдущему, следующие же меридианальные деления идут в очень разнообразных направлениях и делят центральное поле бластодиска на неравныебластомеры. При этом с пятого деления возникают и тангентальные (параллельные поверхности
яйца) плоскости деления, отделяющие мелкие свободные бластомерыот бластомер, соединенных с желтком. Так как такое деление здесь,повторяется многократно, то к концу деления бластомеры, отделяющиеся от желточных бластомер,образуют нижний слой клеток, лежащий значительно ниже, поверхности яйца во впадине, наполненной жидкостью. После обособления бластомер последнего, самого нижнего слоя продолжаетсяеще довольно долго обособлениетаким же путем отдельных клеток,утрачивающих связь с желтком, кпроцесс этот очень постепенно замирает. Ядра, оставшиеся в оченьнебольшом числе в протоплазме поверхностного слоя желтка под центральным полембластодиска, смещаются вместе с остатками протоплазмы в периферическое поле его,так что дно впадины образовано не перибластом, а желтком с очень небольшим количеством протоплазмы (рис. 329, В).
Когда борозды дробления начинаютпроникать из центрального в периферическое поле бластодиска, то они разде  ляют здесь протоплазму на крупные сегменты, которые и при дальнейших своихделениях остаются связанными с желтком, т. е. свободных бластомер от себяне отделяют. Получается здесь такимобразом пласт крупных, медленно делящихся бластомер, связанных с желтком.
ляют здесь протоплазму на крупные сегменты, которые и при дальнейших своихделениях остаются связанными с желтком, т. е. свободных бластомер от себяне отделяют. Получается здесь такимобразом пласт крупных, медленно делящихся бластомер, связанных с желтком.
Таким образом дробление приводит кстадии, на которой центральная частьбластодиска состоит снаружи из свободных мелких бластомер, хотя и округленных, но соединенных друг с другом вдовольно плотный слой, а под ними вполости с жидкостью лежит очень болььшое количество также свободных бластомер; боковые стенки этой полости образованы краем пласта крупных периферических бластомер, и такие же крупныебластомеры, но в очень небольшом числе,образуют и дио полости. Эту стадиюможно назвать бластомерной бластулой.
Бластомерная бластула без деления клеток превращается в эпителиальную, причем бластомеры бластоцеля подходят и присоединяются к наружному слою бластодиска; вследствие этого последний становится еще более плотным и несколько увеличивается в диаметре, клетки же его получают характер эпителия. Эпителий при этом надвигается на периферическое поле бластодиска, причем ложится здесь прямо на его протоплазму и желток.
По внешности бластодиск на стадии бластулы имеет, как и прежде, вид круглого пятна на поверхности желтка; но помимо того, что пятно это стало многоклеточным, и диаметр его несколько увеличился, оно отличается от первоначального зародышевого пятна тем, что состоит из двух частей: центральной, более светлой, благодаря просвечивающей через наружный эпителий жидкости бластоцеля и называемой поэтому светлым или просвечивающим полем, или area pellucida, и периферической, где бластодиск лежит прямо па желтке, и называемой темным полем, или area ораса.
 На этой и на последующих стадиях развития не прекращается процесс деления бластомер в area ораса, но он принимает несколько измененный характер и называется дополнительным дроблением. В этой периферической зоне, с самого начала отставшей в своем дроблении от центральной, продолжается обособление новых бластомер, дальнейшая диференцировка которых идет гораздо проще, чем в центре, да и сам способ их обособления совершенно своеобразен. Здесь лежит довольно массивный пласт протоплазмы, в котором идет деление ядер, не сопровождающееся однако делением протоплазм, а ведущее к образованию синцития, к которому позднее присоединяются редкие ядра и протоплазма из под area pellucida. Некоторые ядра этого краевого синцития вместе с довольно большим участком протоплазмы, в который перед этим вступают зерна желтка, отделяются от синцития в виде крупных клеток. Из них постепенно слагается слой на желтке, распространяющийся по всем радиусам вместе с ростом бластодиска и представляющий собой внезародышевую, или желточную, эптодерму. Прикрывающая ее сверху внезародышевая эктодерма с подстилающей последнюю мезенхимой возникает разрастанием соответственных частей area pellucida. Этот процесс заканчивается на поздних стадиях почти полным обрастанием желтка всеми этими тремя слоями. По характеру клеток желточной энтодермы и по изменениям в них во время развития можно заключить, что желточная энтодерма принимает прямое участие в усвоении желтка, и является функционирующим во время развития органом, но никакого участия в построении кишечника птицы не принимает и при резорбции всего желтка уничтожается.
На этой и на последующих стадиях развития не прекращается процесс деления бластомер в area ораса, но он принимает несколько измененный характер и называется дополнительным дроблением. В этой периферической зоне, с самого начала отставшей в своем дроблении от центральной, продолжается обособление новых бластомер, дальнейшая диференцировка которых идет гораздо проще, чем в центре, да и сам способ их обособления совершенно своеобразен. Здесь лежит довольно массивный пласт протоплазмы, в котором идет деление ядер, не сопровождающееся однако делением протоплазм, а ведущее к образованию синцития, к которому позднее присоединяются редкие ядра и протоплазма из под area pellucida. Некоторые ядра этого краевого синцития вместе с довольно большим участком протоплазмы, в который перед этим вступают зерна желтка, отделяются от синцития в виде крупных клеток. Из них постепенно слагается слой на желтке, распространяющийся по всем радиусам вместе с ростом бластодиска и представляющий собой внезародышевую, или желточную, эптодерму. Прикрывающая ее сверху внезародышевая эктодерма с подстилающей последнюю мезенхимой возникает разрастанием соответственных частей area pellucida. Этот процесс заканчивается на поздних стадиях почти полным обрастанием желтка всеми этими тремя слоями. По характеру клеток желточной энтодермы и по изменениям в них во время развития можно заключить, что желточная энтодерма принимает прямое участие в усвоении желтка, и является функционирующим во время развития органом, но никакого участия в построении кишечника птицы не принимает и при резорбции всего желтка уничтожается.
Гаструляция. Возвращаясь к стадии эпителиальной бластулы, проследим дальнейшие происходящие в ней изменения.
Прежде всего в ее бластоцеле появляется второй эпителиальный слой, который слагается из очень плоских клеток, подстилает всю area pellucida (рис. 330) и представляет собой кишечную энтодерму. Слой этот возникает из заднего края бластодиска, где на этой стадии имеется перерыв в area ораса, так как бластоцель area pellucida граничит здесь не с плазмой area ораса, а непосредственно с желтком; наружный эпителий бластодиска утончен около этого места до одного слоя клеток. Через этот задний край происходит подворачивание и рост слоя клеток в бластоцель, которыедают тонкий листок кишечной энтодермы (рис. 331). Таким способом происходитначальная гаструляция, которая имеет большое сходство с таковой Anamnia, таккак место подворачивания или бластопор лежит па этой стадии на краю бластодиска.Кишечная энтодермане соприкасается с желтком и не функционирует у зародыша.Однако таким путем возникает только энтодерма, притом очень тонная и неспособнаядать какие-либо органы осевого комплекса, возникающего при гаструляции позвоночных.
После образования кишечной энтодермы, которая своими краями соединяется с желточной энтодермой area ораса, процесс гаструляции прерывается, и в это время происходит перемещение материала, прежде всего выражающееся в том, что позади закрывшегося теперь бластопора замыкается кольцо area ораса, так что бластопор оказываетсятеперь на заднем краю area pelluсida и удален от заднего края бластодиска. На местезакрывшегося бластопора собираются теперь клетки, которые приходят с боков заднего края area pelluсida на место ушедшего в бластоцель материала кишечной энтодермы и представляют собой материал мезодермы, образующий серповидное утолщение в задней части area pelluсida (рис. 332, А).
 Area pelluсida при этом несколько вытягивается в длину и принимает расширеннуювпереди грушевидную форму. Ее заднее утолщение вытягивается прямо вперед в виде продольной утолщенной полостей, называемой первичной полоской (рис. 332, В, 3); повидимому, ее вытягивание не естьпередвижение клеток утолщения вперед или бывает таковым только отчасти, а является результатом вытягивания areapelluсida назад. После достижения первичной полоской определенной длины эпителий areapelluсida вырастает позади от еезаднего конца, так что полоскалежит теперь на некоторомрасстоянии впереди заднего края area pellueida и еще дальше от заднего края area ораса.Эти перемещения материалапервичной полоски выяснены при помощи метода прижизненной маркировки бластодиска на различных стадиях. Впереди первичная полосказаканчивается небольшим круглым расширением, называемым гензеновским, или первичным узелком (рис. 332, 4). Полоска очень отчетливо видна на прозрачном фоне areapelluсida благодаря тому, что по всей ее длине идет интенсивная миграция клеток в бластоцель, так что иод ней лежит густое и многослойное скопление этих клеток (рис. 333).Так как миграция идет по срединной линии полоски, то по этой линии в ней заметнапродольная бороздка. Миграция идет и в геизеновском узелке, а бороздка здесь заканчивается круглой ямкой, направленной своим дном вперед.
Area pelluсida при этом несколько вытягивается в длину и принимает расширеннуювпереди грушевидную форму. Ее заднее утолщение вытягивается прямо вперед в виде продольной утолщенной полостей, называемой первичной полоской (рис. 332, В, 3); повидимому, ее вытягивание не естьпередвижение клеток утолщения вперед или бывает таковым только отчасти, а является результатом вытягивания areapelluсida назад. После достижения первичной полоской определенной длины эпителий areapelluсida вырастает позади от еезаднего конца, так что полоскалежит теперь на некоторомрасстоянии впереди заднего края area pellueida и еще дальше от заднего края area ораса.Эти перемещения материалапервичной полоски выяснены при помощи метода прижизненной маркировки бластодиска на различных стадиях. Впереди первичная полосказаканчивается небольшим круглым расширением, называемым гензеновским, или первичным узелком (рис. 332, 4). Полоска очень отчетливо видна на прозрачном фоне areapelluсida благодаря тому, что по всей ее длине идет интенсивная миграция клеток в бластоцель, так что иод ней лежит густое и многослойное скопление этих клеток (рис. 333).Так как миграция идет по срединной линии полоски, то по этой линии в ней заметнапродольная бороздка. Миграция идет и в геизеновском узелке, а бороздка здесь заканчивается круглой ямкой, направленной своим дном вперед.
 Мигрирующие из первичной полоски мезодермальные клетки заполняют в бластоцеле все пространство между ней и кишечной энтодермой и расходятся в стороны болеетонким слоем между эктодермой и кишечной энтодермой, а затем продолжения этихбоковых пластинок ее проникают между эктодермой и желточной энтодермой area ораса.В области гензеновского узелка часть мезодермальных клеток продвигается вперед ив стороны от него.
Мигрирующие из первичной полоски мезодермальные клетки заполняют в бластоцеле все пространство между ней и кишечной энтодермой и расходятся в стороны болеетонким слоем между эктодермой и кишечной энтодермой, а затем продолжения этихбоковых пластинок ее проникают между эктодермой и желточной энтодермой area ораса.В области гензеновского узелка часть мезодермальных клеток продвигается вперед ив стороны от него.
Вслед за сформированием всей первичной полоски с первичной бороздкой и гензеновским узелком полоска начинает медленно укорачиваться вследствие перемещениягензеновского узелка по направлению к заднему концу ее. При этом мезодерма, возникшая из передней части полоски, остается на месте, и, по мере сдвигания гензеновскогоузелка кзади, впереди него остается срединный тяж, кажущийся продолжением первичной полоски, но менее ясно очерченный, чем эта последняя (рис. 332, С). Тяж, появляющийся впереди гензеновского узелка, представляет собою только осевую утолщеннуючасть находящейся здесь мезодермы, но по сторонам он, как п в области первичной полоски, продолжается в боковые, более тонкие мезодермальные пластинки, краевыеклетки которых проникают и под эктодерму area ораса. Этот осевой тяж мезодермы впереди гензеновского узелка получил наименование головного отростка.
Головной отросток состоит из более плотно расположенных клеток, чем первичнаяполоска, и теснее, чем мезодерма последней, прилегает к кишечной энтодерме(рис. 333, В, 5); вместе с тем головной отросток отодвинут от эктодермы и отделен от нееучастком бластоцеля, никакоймиграции из наружного слоя внем нет, и потому он менее ясно виден с поверхности. Дальнейшая диференцировка головного отростка показывает, что в состав его входит не только мезодерма, по и хорда, которая занимает его узкую осевую часть,тогда как утолщенный пласт посторонам хорды дает миотомы,а боковые, более тонкие слои представляют собой зачаток боковых пластинок или спланхнотомов. При этом эктодерма,значительно утолщенная над головным отростком, так и впереди него, образует нервные валики, сначала раздвинутые в стороны и позднее сближающиеся друг с другом в нервнуютрубку. Все эти проявленияорганизации и диференцировки происходят только в области головного отростка, т. е. в той части первичной полоски, котораяпри отодвигании гензеновскогоузелка кзади оказалась впередиэтого последнего.
 Довольно большое число сделанных различными авторами экспериментальных исследований дают не всегда одинаковое толкование процесса образования головного отростка. Мы приведем здесь то из них, которое объясняет процесс в согласии с данными о возникновенииорганизации в эмбриональномразвитии других позвоночных;это толкование дано двумя различными авторами (Ветцель и Грэпер), которые пришли к нему; один методом маркировки, другой — методом прижизненной кинематографической съемки развития зародыша.
Довольно большое число сделанных различными авторами экспериментальных исследований дают не всегда одинаковое толкование процесса образования головного отростка. Мы приведем здесь то из них, которое объясняет процесс в согласии с данными о возникновенииорганизации в эмбриональномразвитии других позвоночных;это толкование дано двумя различными авторами (Ветцель и Грэпер), которые пришли к нему; один методом маркировки, другой — методом прижизненной кинематографической съемки развития зародыша.
Так как в первичной полоске мы находим только неорганизованный материал, тоорганизация наступает впереди гензеновского узелка потому, что через последнийвносится недостающий зачаток, без которого организация осевых зачатков не наступаети у других позвоночных. Таким зачатком является зачаток хорды. Самое же передвижение гензеновского узелка кзади аналогично нарастанию верхней губы бластопора,создающему контакт между всеми осевыми зачатками хордовых. При этом нарастанииверхняя сторона образуемой верхней губой складки дает у всех хордовых нервную пластинку, а нижняя — хордальную пластинку, и обе они удлиняются по мере удлиненияголовного отростка, сопровождающегося укорачиванием первичной полоски. Клетки,возникающие иэ первичной полоски, представляют собою мсзодермальный материал,как это видно из их дальнейшего развития. Во время прохождения по этому материалугензеновского узелка или верхней губы бластопора в неорганизованный мезодер мальныйматериал вступает хордальная пластинка, которая тесно соединяется с кишечной энтодермой и притягивает к себе части мезодермы, образующей по сторонам ее более плотныеутолщения. Так как на поверхности бластодиска на это место надвигается нервнаяпластинка или нервные валики, то таким образом создается весь характерный дляChordata осевой или хордальный комплекс зачатков.
Следовательно, образование первичной полоски и возникновение из нее головногоотростка путем передвижения гензеновского узелка есть также процесс гаструляции, каки закладка кишечной энтодермы; у птиц, следовательно, гаструляция разделена на двефазы, отграниченные друг от друга и во времени и тем, что в промежутке между нимипроисходит перемещение бластопора. При образовании кишечной энтодермы он находится на за днем краю area pellucida, но затем отходит от него вперед и вытягивается по продольной оси зародыша, образуя первичную полоску и гензеновский узелок; при этом вовторой фазе отверстие бластопора сохраняется только в передней его части,т. е. в гензеновском узелке, на протяжении же первичной полоски он замкнут, и миграция клеток в бластоцель идет выклиниванием отдельных клеток из него. В отличие от Anamnia боковые края бластопора сближены друг с другом в продольную полоску, так как у Anamnia они раздвинуты в кольцо эктодермальным материалом,у птиц же эптодермальныйматериал в первой фазе гаструляции уходит в бластоцель и не влияет нарасположение зачатков вовторой фазе.
Разобщенность закладки кишечной энтодермы от остальных осевых зачатков представляет интерес еще в другом отношении. У Anamnia при гаструляции все осевыезачатки возникают одновременно и образуют совместно стенку гастральной полости,что дало повод считать, что хорда и мезодерма возникают энтероцельно из стенки единого зачатка — первичной энтодермы. Но у птиц и прочих Anmiota отношения этихзачатков, при которых энтодерма закладывается раньше и отдельно от других зачатков,и всетаки во второй фазе гаструляции вступает в тесный контакт с хордой и мезодермой,показывают, что такая связь их между собой и у Anamnia и у Amniota является лишьследствием необходимости для начала диференцировки создания временного контактамежду всеми этими зачатками.
Из всех экспериментальных исследований над развитием птиц можно кроме тогозаключить, что в области первичной полоски и головного отростка возникает не всянервная система. Если до начала образования головного отростка сделать на живомбластодиске поперечный надрез сейчас же впереди гензеновского узелка, то рана растягивается в виде крупного отверстия в блаетодиске, и впереди переднего ее края возникают через день передний и средний мозговые пузыри, а позади ее заднего края — нервная трубка не развивается (рис. 334, А); другими словами, гензеновский узелок поположению своему до образования головного отростка отвечает переднему концу хорды,а прехордальный отдел нервной трубки возникает впереди и гензеновского узелка ихорды, связанной с узелком по своему возникновению. При полном удалении гензеновского узелка на той же стадии тело зародыша не организуется, но передний и средниймозговые пузыри развиваются нормально(рис. 334, В). Мезодерма этой прехордальной области головы возникает частью изпродвигающихся вперед клеток из первичной полоски, частью образуется на местеиз прехордальной пластинки кишечника.Если же сделать надрез впереди гензеновского узелка во время образования головного отростка, т. е. перерезать этот последний, то впереди раны развивается тем больший передний участок всего тела, чем позднее была стадия, на которой была сделанаоперация, и чем длиннее, был головной отросток (рис. 334, С). При этомнад отверстиемраны от ее переднего края вырастает отросток хорды, что говорит о связи хорды сголовным отростком и с верхней губой бластопора (рис. 334, В), равно как и то, чтопри пересадке гензеновского узелка в какую-нибудь часть area pellucida он даетздесь только хорду.
 С возникновением контакта между всемиосевыми зачатками у птиц создается та же группировка и те же их взаимоотношения,какие характеризуют всех позвоночных иланцетника. Затем все эти зачатки обособляются друг от друга вследствие формированияиз хордальной пластинки хордального тяжа,и сохраняют способность к взаимодействиюпри своем дальнейшем формировании. Нервная пластинка в присутствии хорды замыкается в нервную трубку, ближайшие кхорде участки мезодермы диференцируютсяв миотомы, а кишечная энтодерма начинаетобразовывать кишечную трубку с ее отделами и придаточными органами.
С возникновением контакта между всемиосевыми зачатками у птиц создается та же группировка и те же их взаимоотношения,какие характеризуют всех позвоночных иланцетника. Затем все эти зачатки обособляются друг от друга вследствие формированияиз хордальной пластинки хордального тяжа,и сохраняют способность к взаимодействиюпри своем дальнейшем формировании. Нервная пластинка в присутствии хорды замыкается в нервную трубку, ближайшие кхорде участки мезодермы диференцируютсяв миотомы, а кишечная энтодерма начинаетобразовывать кишечную трубку с ее отделами и придаточными органами.
По мере перемещения гензеновскогоузелка назад головной отросток удлиняется,и в то время, как диференцируются зачаткив задних его участках, передняя часть зародыша уже. довольно далеко продвигаетсяв своей диференцировке как общей формытела, так и отдельных зачатков.
Одна из наиболее характерных особенностей гаструляции птиц и всех Amniotaсостоит в том, что в то время, как у Anamnia с дискоидальным дроблением бластопор находится всегда на краю диска, у Amniotaон появляется на краю area pellucida и, следовательно, отделен от края бластодискакольцом area ораса, а позднее смещается в самое area pellucida. Поэтому край бластодиска или край area ораса становится исключительно краем обрастания, вследствие чегообрастание идет равномерно по всем меридианам желтка.
В течение гаструляции и позднее, при дальнейшем обрастании желтка бластодермой, на желток надвигается описанным выше образом желточная энтодерма, а также эктодерма от краев area pellucida и мезенхима, возникающая из краевой части мезодермы первичной полоски, которая в виде клеток проникает в area ораса. Из этой мезенхимы еще до окончания гаструляции возникают кровяные островки, которые дают сеть желточных кровеносных сосудов; с этого времени area ораса именуется сосудистым полем. На более поздпих стадиях капилляры, возникшие раньше и лежащие ближе к зародышу, становятся более крупными желточными артериями и венами; а на краю разрастающегося сосудистого поля возникает из мезенхимы камбиальный пояс, непрерывно продуцирующий новые кровяные островки и новые мелкие капилляры.
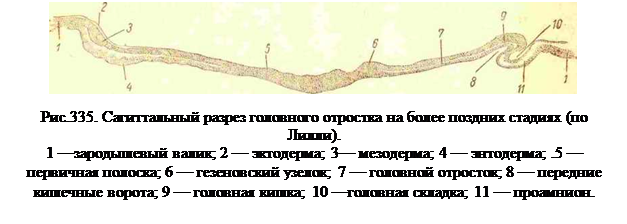 Лежащие впереди от головного отростка нервные валики, дающие передний и средний мозговые пузыри, вырастают и в ширину и в вышину сильней остальных частей валиков и кроме того растут своим передним краем вперед, наклоняясь над лежащей впереди бластодермой (рис. 335). Так как здесь нервные валики приподнимают с собой и эктодерму, то приподнимается над бластодермой вся передняя половина головы, и этот свободный конец отграничивается от бластодермы головной складкой (рис. 335,10). За нею постепенно спереди назад приподнимаются позади лежащие части зародыша. Приподнимание тела зародыша вызывается, однако не ростом внутренних органов, а особым процессом, напоминающим поднимание зародыша у селяхий и связанным в данном случае с образованием амниона, о чем речь будет несколько дальше.В приподнимающуюся часть зародыша втягивается кишечная энтодерма, которая образует внутри зародыша продольную карманообразную складку. При образовании у зародыша головной складки передни л конец кишечной складки принимает вид трубчатого кармана, называемого головной кишкой поткрывающегося в эктодермальную полость головными кишечными воротами (рис. 335,8). По мере дальнейшего обособления переднего конца зародыша от внезародышевой бластодермы эти кишечные ворота смещаются кзади, а трубчатая часть кишечной энтодермы все более и более удлиняется.
Лежащие впереди от головного отростка нервные валики, дающие передний и средний мозговые пузыри, вырастают и в ширину и в вышину сильней остальных частей валиков и кроме того растут своим передним краем вперед, наклоняясь над лежащей впереди бластодермой (рис. 335). Так как здесь нервные валики приподнимают с собой и эктодерму, то приподнимается над бластодермой вся передняя половина головы, и этот свободный конец отграничивается от бластодермы головной складкой (рис. 335,10). За нею постепенно спереди назад приподнимаются позади лежащие части зародыша. Приподнимание тела зародыша вызывается, однако не ростом внутренних органов, а особым процессом, напоминающим поднимание зародыша у селяхий и связанным в данном случае с образованием амниона, о чем речь будет несколько дальше.В приподнимающуюся часть зародыша втягивается кишечная энтодерма, которая образует внутри зародыша продольную карманообразную складку. При образовании у зародыша головной складки передни л конец кишечной складки принимает вид трубчатого кармана, называемого головной кишкой поткрывающегося в эктодермальную полость головными кишечными воротами (рис. 335,8). По мере дальнейшего обособления переднего конца зародыша от внезародышевой бластодермы эти кишечные ворота смещаются кзади, а трубчатая часть кишечной энтодермы все более и более удлиняется.
В процессе своего перемещения кзади гашеповскнй узелок, однако не достигает заднего конца первичной полоски, так что после окончан ья второй фазы гаструляции позади его остается недеференцированный участок, который представляет собой так называемый туловищно-хвостовой зачаток, гомологичный хвостовой почке Ananmia. Этот зачаток на данной стадии сравнительно мало выступает на поверхности, и его клеточная масеа вдавлена вниз по направлению к желтку, так что сзади и с боков она ограничена экзоцеломом (рис. 337). Сплаихноплевра экзоцелома совместно с прилегающей к ней кишечной энтодермой начинает впячиваться вперед под ткань, отделяя ее от внезародышевой энтодермы и епланхноплевры желтка. Вследствие этого кишечная энтодерма образует складку, направленную назад и называемую хвостовыми кишечными воротами (pис. 337, А, 4). После такого внутреннего обособления туловищно- хвостового зачатка начинается образование входящей складки еоматоплевры экзоцелома и эктодермы, которые значительно глубже впячиваются под нее, как хвостовая складка амниона, и отделяют туловищно-хвостовой зачаток от внезародышевой эктодермы.
 Образование зародышевых оболочек. Головная складка возникает гораздо раньше хвостовой, тем не менее образование и той и другой суть части одного и того же процесса, именно образования амниона. Способ его возникновения описан во вступлении к описанию развития Arnniota, здесь же отметим только, что его образование начинается с головного конца, причем место его появления можно указать еще до фактического его образования. К этому времени мезенхима распространяется подо всем бластодиском, но впереди гол
Образование зародышевых оболочек. Головная складка возникает гораздо раньше хвостовой, тем не менее образование и той и другой суть части одного и того же процесса, именно образования амниона. Способ его возникновения описан во вступлении к описанию развития Arnniota, здесь же отметим только, что его образование начинается с головного конца, причем место его появления можно указать еще до фактического его образования. К этому времени мезенхима распространяется подо всем бластодиском, но впереди гол