Фотосинтез – это процесс образ-я органич-х в-в из СО2 и Н2О при помощи пигментов зеленого листа и энергии солнечного света. Ф-з – это процесс преобразования энергии солнечного света в энергию хим-х связей орг-х соед-й.
 6СО2 + 6Н2О свет, пигмент С6Н12О6 + 6О2.
6СО2 + 6Н2О свет, пигмент С6Н12О6 + 6О2.
Важнейшим компонентом аппарата фотосинтеза явл-ся пигменты. Пигменты относ-ся к 3 классам в-в: хлорофиллам, фикобилинам и каротиноидам. Хлорофилл – пигмент зеленого цвета. Это сложный эфир дикарбоновой кислоты хлорофиллина и 2-х спиртов: метилового и фитола. У высших раст. и водорослей обнаружены хлорофиллы а, b, c, d. Все фотосинтез-ие раст., вкл. все группы водорослей и цианобактерии, содержат хлор-л а. Хлорофилл b – у высших раст. и зелен. водорослей и эвгленовых. У бурых и диатомовых вод-лей вместо хлор-ла b присутствует хлор-л с, а у многих красных вод-лей – хлор-л d.
Каротиноиды – пигменты желтого, оранжевого, красного цвета – присутствуют в хлоропластах всех раст. Вкл. в себя 3 типа соединений: каратины, ксантофиллы и каратиноидные к-ты (придукт окисления первых 2-х групп). Фолиевые соединения.
Фикобилины – это пигменты, хар-ные с-з водорослям, цианобактериям и красным водорослям. Представители – фикоэритробилины и фикоцианобилины.
Пигменты в мембранах хлоропластов (это мембранный гран и тилакоиды стромы) образуют светообразующий комплекс, кот. состоит из молекул - антенн и молекулы – ловушки (ССК). Молекулами – ловушками м.б. хлор-л а – Р700, и Р680. мол-ми – антеннами м.б. хлор-л а, хлор-л b, каротиноиды.
Фотофизич-ий этап фотос-за. Важнейшей особенностью фотос-за явл. то, что он протекает с использованием энергии солнечного света. Благодаря фотосистемам (ФС1 –Р700 - и ФС2 –Р680) = ССК+ ЭТЦ при освещении синим светом е- переходят на орбиталь с более высоким энергетическим уровнем – S2 (синглетное состояние), где нах-ся непродолжительное время 10-13 – 10-14 сек., после чего они перемещ-ся в сторону ядра – на S1, а выделяемая при этом энергия тратится на образование теплоты, флюоресценции и выполнение фотохимич-ой работы. На этот же уровень е- попадают при освещении красным светом. На S1 е- так же нах-ся непродолжительное время 10-9 сек., откуда они возвращ-ся в S0 (если не измен-ся спин е-), либо в триплетное состояние Т1 (если один е- изменяет спин). На Т1 уровне е- нах-ся самое продолжительное время – 10-4 сек., там он либо успевает подхватиться первичным белком акцептором е- и тогда запускается в ЭТЦ фотос-за, либо успевает поменять свой спин и возвращ-ся на S0 уровень, причем выделяемая энергия так же использ-ся на фотохим-ую работу.
Перенос е- по цепи переносчиков вкл. ряд окислительно-восстановит-х реакций, в ходе кот. выдел-ся энергия (она м.б. использована для синтеза АТФ) – фотосинтетическое фосфорилирование. Различают 2 типа ф-го фосфор-я: циклическое и нециклическое.

 Ц. фосфор-ние протекает как на мембранах гран, так и в тилакоидах стром. Наиболее древний путь ф-за, в кот. уч-ет только ФС1, в кот. мол. Р700 явл. как донором, так и акцептором е-. свет падает на хлоропласт раздражая мембрану = возбуждение Р700 - ее е- переходят на удаленную орбиталь и подхват-ся первичным белком акцептором е- переходит на белок-переносчик ферредок-син цитохром b цитохром f возвращ-ся в Р700. М/ду цитохромами е- движ-ся в противоположном направлении по мембране, поэтому он выделяет энергию, кот. тратится на синтез АТФ. Хар-рен пуриновым и зелен. бактериям, высшим раст.
Ц. фосфор-ние протекает как на мембранах гран, так и в тилакоидах стром. Наиболее древний путь ф-за, в кот. уч-ет только ФС1, в кот. мол. Р700 явл. как донором, так и акцептором е-. свет падает на хлоропласт раздражая мембрану = возбуждение Р700 - ее е- переходят на удаленную орбиталь и подхват-ся первичным белком акцептором е- переходит на белок-переносчик ферредок-син цитохром b цитохром f возвращ-ся в Р700. М/ду цитохромами е- движ-ся в противоположном направлении по мембране, поэтому он выделяет энергию, кот. тратится на синтез АТФ. Хар-рен пуриновым и зелен. бактериям, высшим раст.
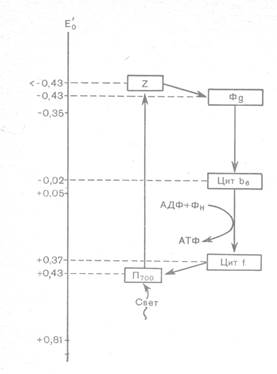
 Нецикл. Фосфор-ие протекает только на мембранах гран. Свет раздражает мембрану, при этом возбужд-ся Р700 и ее е- перенос-ся на акцептор Z и далее по системе переносчиков е- попадает на ферредоксин флавопротеид на наруж. сторону мембраны, заряжая ее ´-´. Белки в ЭТЦ располаг-ся по увеличению окислительно-восстан-го потенциала. Р700 отдав е- стан-ся ´+´ заряженной, в ней образ-ся «электронная дырка». Недостающий е- Р700 получает от ФС2, Р680. Последняя так же возбужд-ся светом и ее е- подхватывается акцептором Z е- передается на цитохром b пластохинон цитохром f. При движении от пластохинона к цитохрому f е- выделяет энергию, благодаря кот. синтезир-ся АТФ. С цитохрома f пластоцианин, а затем е- заполняет «электронную дырку» в Р700. на данном этапе окисленной явл. Р680. Недостающие е- она забирает от воды, кот. нах-ся внутри мембраны грана.
Нецикл. Фосфор-ие протекает только на мембранах гран. Свет раздражает мембрану, при этом возбужд-ся Р700 и ее е- перенос-ся на акцептор Z и далее по системе переносчиков е- попадает на ферредоксин флавопротеид на наруж. сторону мембраны, заряжая ее ´-´. Белки в ЭТЦ располаг-ся по увеличению окислительно-восстан-го потенциала. Р700 отдав е- стан-ся ´+´ заряженной, в ней образ-ся «электронная дырка». Недостающий е- Р700 получает от ФС2, Р680. Последняя так же возбужд-ся светом и ее е- подхватывается акцептором Z е- передается на цитохром b пластохинон цитохром f. При движении от пластохинона к цитохрому f е- выделяет энергию, благодаря кот. синтезир-ся АТФ. С цитохрома f пластоцианин, а затем е- заполняет «электронную дырку» в Р700. на данном этапе окисленной явл. Р680. Недостающие е- она забирает от воды, кот. нах-ся внутри мембраны грана.
 2Н2О свет, Р 680 4Н+ + 4е- + О2 (р-я Хилла, р-я фотолиза). О2 направл-ся в атмосферу, Р680 восстан-ся е-, в переносе кот. уч-ют ионы Mn. Протоны, сосредоточенны на внутр. стороне мемб. и заряжают ее ´+´. Т.о. на мемб. Устан-ся разность потенциалов (электрохим-ий потенциал). Мембрана прошнурована ферментом АТФ-синтетаза.. Протоны способны перемещ-ся с внутр. стороны мемб. на наружную, причем происходит разрядка мемб. с выделением энергии, кот. тратится на синтез АТФ (2-й продукт свет. стадии). Часть протонов вместе с е- восстанавлив-т дегидрогеназу с коферментом НАД+ до НАДН2 (3- продукт с.ст.), т.о. нециклич. фосфор-ие заключ-ся в переносе воды с внутр. стороны мемб. на наруж. = 1 О2 уходит в атмосферу, АТФ и НАДН2 идут на реакции темновой стадии.
2Н2О свет, Р 680 4Н+ + 4е- + О2 (р-я Хилла, р-я фотолиза). О2 направл-ся в атмосферу, Р680 восстан-ся е-, в переносе кот. уч-ют ионы Mn. Протоны, сосредоточенны на внутр. стороне мемб. и заряжают ее ´+´. Т.о. на мемб. Устан-ся разность потенциалов (электрохим-ий потенциал). Мембрана прошнурована ферментом АТФ-синтетаза.. Протоны способны перемещ-ся с внутр. стороны мемб. на наружную, причем происходит разрядка мемб. с выделением энергии, кот. тратится на синтез АТФ (2-й продукт свет. стадии). Часть протонов вместе с е- восстанавлив-т дегидрогеназу с коферментом НАД+ до НАДН2 (3- продукт с.ст.), т.о. нециклич. фосфор-ие заключ-ся в переносе воды с внутр. стороны мемб. на наруж. = 1 О2 уходит в атмосферу, АТФ и НАДН2 идут на реакции темновой стадии.
| 
| Фд | 
|
 ФП ФП
| 
| наруж. стор. Мем. ´-´ |  свет свет
| ||

| ФС1 | П 680 | |||||||
 П 700 П 700
| Цит b | 
| |||||||
 свет свет
| ПЦ | 
|  Цит f Цит f
| 
|  Z Z
| ||||

| ПХ | ФС2 | |||||||
| АДФ + Фн |
Схема нециклич. транспорта электронов при ф-зе:
Фд – ферредоксин; ФП – флавопротеид; ПХ – пластохинон;
ПЦ – пластоцианин; Z – неиденфицированный переносчик.
Хар-рен для цианобактерий, некот. водорослей и всех высших растений.
Темновая стадия фотосинтеза (цикл Кальвина, С3 - путь) протекает в строме хлоропласта. Не требует освещения, но нужд-ся в продуктах световой стадии – АТФ и НАДН2.

Цикл Кальвина.
В строме хлоропласта есть 5-углеродный сахар – рибулезадифосфат (РДФ). С помощью фермента рибулезадифосфат-карбоксилаза он присоед-т СО2 = нестойкое 6-углеродное соединение, кот. распадается до фосфоглицериновой к-ты (ФГК), кот. активируется АТФ = дифосфо-глицериновую к-ту и восстанавл-ся НАДН2 до фосфоглицеринового альдегида (ФГА). Часть ФГА идет на регенерацию ребулезодифосфата, причем тратится еще 6 мол-л АТФ, из 2-х оставшихся мол-л ФГА одна изомеризуется в диокси-ацетонфосфат (ДОАФ). Взаимодействуя м/ду собой эти 2 мол-лы = фруктозадифосфат (ФдиФ), кот. отщепляя 1 фосфатную гр. = фруктозамоно-фосфат (ФМФ). ФМФ изомеризуется в глюкозамонофосфат (ГМФ), кот. отщепляет фосфатную гр. = глюкоза.
 6СО2 + 12НАДН2 + 18 АТФ С6Н12О6 + 12НАД+ + 18АТФ + 18 Фн+ + 6Н2О
6СО2 + 12НАДН2 + 18 АТФ С6Н12О6 + 12НАД+ + 18АТФ + 18 Фн+ + 6Н2О
 из световой стадии
из световой стадии
В темновой стадии фот-за на ассимиляцию СО2 использ-ся не весь РДФ, а только часть, а вторая часть уходит на фотодыхание (это процесс усиленного расщепления углеводов и усиленного потребление О2 на свету, а следовательно усиленное выделение СО2.
Для некоторых южных злаков (просо, кукуруза, сахарный тростник и др.) хар-рен  цикл
цикл
Хетча-Слэка-Карпилова (С4- путь)
Данный процесс не заменяет цикла Кальвина, а играет по отношению к нему вспомогательную роль. Поскольку основными метаболитами этого цикла оказались четырех - углеродные соединения, его еще называют С4-путь, а растения, — С4-растениями. В цитоплазме клеток мезофилла осуществляются реакции цикла Хэтча — Слэка — Карпилова, а в хлоропластах клеток обкладки — преимущественно реакции цикла Кальвина. Согласованное функционирование этих двух циклов послужило основанием назвать цикл Хетча — Слэка — Карпилова кооперативным фотосинтезом. Он осуществляется следующи образом
Этот путь ф-за разграничен в пространстве, т.е. протекает в 2-х кл. С4 – раст. имеют 2 типа хлоропластов – одни мелкие гранальные, нах-ся в кл. мезофилла листа (зелен. кл.), др. агранальные, нах. в клетках обкладки и окружают жилки. В кл. мезофилла происходит фосфорилирование пировиноградной к-ты (ПВК) до фосфоэнолпируат, кот. карбоксилир-ся образуя ЩУК, кот. восстан-ся НАДН2 до яблочной кислоты (ЯК). ЯК диффундирует в кл. обкладки, там окисл-ся с образованием НАДН2, СО2 и ПВК(ЩУК. ЩУК декарбоксилир-ясь отдает СО2, связ-ся с РДФ и ЩУК превр-ся в ПВК). СО2 там же вступает в цикл Кальвина, а ПВК возвр-ся в кл. мезофилла.
У толстянковых (кактусов, очитков) своеобразный путь ф-за - САМ-путь. САМ- путь по химизму схож с С4- путем, только реакции разграничены не в пространстве, а во времени. Связывание органич-ми к-тами СО2 происходит ночью при открытых устьицах, а световая фаза фотосинтеза и синтез орг-го в-ва - днем при закрытых устьицах.
8. Дыхание как энергетический процесс, его механизмы и значение в жизни растительного организма. Пути дыхательного обмена и эволюция дыхания.
В природе существуют два основных процесса, в ходе которых энергия солнечного света, запасенная в органическом веществе, высвобождается — это дыхание и брожение. Дыхание — это аэробный окислительно-восстановит-й энергетич-й процесс, при кот. органич-е в-ва, богатые энергией окисляются до бедных энергией СО2, Н2О
Суммарное уравнение реакции д-я: C6H12О6 + 6О2 = 6СО2 + 6Н2О+ Е кДж
Брожение — это анаэробный окислительно-восстановит-й процесс, при кот. орг-ие в-ва окисл-ся до орг-х в-в еще богатых энергией.