Рассмотренные формы пространственной дифференциации представляют собой механизмы увеличения дисперсности в распределении популяции по территории. Их основное адаптивное значение заключается в снижении конкуренции за ресурсы. Но, как уже говорилось, «центробежный» процесс пространственной дифференциации не может быть бесконечным – этому противостоит задача интеграции особей в пространстве, без чего не могут быть реализованы важнейшие общепопуляционные функции: воспроизводство, регуляция плотности населения, общепопуляционная реакция на внешние условия, взаимоотношения с популяциями других видов. Поэтому в общей системе популяционных адаптаций механизмы, определяющие устойчивое поддержание контактов между особями, их интеграцию в единую функциональную систему, имеют важнейшее биологическое значение. Эти механизмы действуют «центростремительно», у территориальных животных как бы уравновешивая действие пространственного разобщения в создании суммарной пространственно-этологической структуры, в которой достаточно низкий уровень конкуренции сочетается с достаточно высокой контактностью, обеспечивающей бесперебойное функционирование популяции. У стадных животных поддержание устойчивых информативных контактов не только предоставляет условия сохранения адаптивного построения стада (стаи), но и лежит в основе поддержания целостности стада как функциональной популяционной системы.
Поддержание информационных контактов. В основе механизмов интеграции лежат два взаимосвязанных процесса: непрерывная информация о месте локализации отдельных особей и их группировок и наследственно детерминированный стереотип поведения, стимулирующего животное к поиску и поддержанию контактов о себе подобными.
Информация о присутствии особей своего вида обширна и многообразна. В нее входят как «личные сигналы», воспринимаемые визуально, на слух и по запаху, так и различные «попутные» формы информации: следы, тропы, поеди; норы, гнезда, фекалии и т. п. У некоторых рыб в качестве такой информации могут выступать электрические разряды, у ряда видов беспозвоночных – механические колебания воды. Немалое значение имеют все формы обозначения занятой территории, попутно с основной функцией несущие информацию о присутствии особей своего вида, пригодности данной территории для жизни. К этому добавляются специальные формы общения – такие, как сигналы об опасности, половые призывы, реакции испуга и т. п. Вся сумма информации в виде различных сигналов, характерных изменений рельефа и растительности, иных следов жизнедеятельности образует в местах постоянного обитания животных биологическое сигнальное поле (Н.П. Наумов, 1973, 1975, 1977), которое служит мощным стимулом реализации интеграционного поведения.
На сумму информации животные реагируют проявлением специфического поведения. Показано, например, что большие песчанки охотнее селятся на колониях того же вида (даже пустующих), чем в аналогичных местах по соседству; их привлекают норы и измененный рельеф вокруг них. В колониях северных качурок новые особи охотнее селятся вблизи от загнездившихся ранее, ориентируясь на вид гнездовых нор и голоса птиц; сооружение искусственных нор и проигрывание записанных на магнитофон криков успешно способствовали формированию таких колоний (R. Podo1sky, S. Кress, 1989).
Подобное явление характерно для многих других животных. Так, в лабораторных условиях ящерицы Anolis aиratus активно стремятся присоединиться к особям своего вида. Ориентация идёт, по-видимому, на основе зрительной рецепции: при отсутствии особей своего вида эти ящерицы присоединялись к внешне похожим особям близкого вида А. tropidogaster.
У ряда видов морских беспозвоночных (морские уточки, медузы и др.) подвижные личинки оседают и успешно развиваются преимущественно в тех местах, где уже прикрепились другие особи; так рассеянные в толще воды личинки формируют компактные колонии прикреплённых форм. Более подробные данные по мидиям показывают, что наиболее эффективное оседание идёт не в центре колонии, а по периферии её; такое поведение личинок регулируется, вероятно, через концентрацию метаболитов в воде.
Икрометание ряда видов рыб определяется местом расположения уже отложенных кладок. В опытах с Cichlasoma nigrifsciatum выяснено, что если место откладки первой порции икры формируется случайным образом, то последующие кладки с высокой степенью достоверности определяются местом ранее отложенной икры (Р. Weber, S. Weber, 1971). То же известно для других видов рыб, а также для амфибий. Так, в полевых опытах с лесной лягушкой Rana sylvatica кладку икры перенесли в пруд, в котором икрометание еще не началось. Через несколько дней все отложенные в этом пруду кладки концентрировались в непосредственной близости от интродуцированной икры (R. Howard, 1980).
Известно, что головастики бесхвостых амфибий образуют скопления. В экспериментах с жабой Bufo americanus было показано, что они формируются не случайно: при запуске в общий аквариум головастиков разных кладок они вскоре группировались по «родственному» принципу, образуя скопления головастиков одной кладки. Во всех этих случаях интеграция особей скорее всего определяется химическими стимулами.
Давно известно, что в годы депрессии численности грызунов зверьки распределены группами. которые могут располагаться далеко одна от другой, сохраняя внутри себя характерные для вида формы расположения участков и взаимодействия особей. Образованию таких групп предшествует период повышенной подвижности, который можно характеризовать как «поиск контактов». Так, в Московской области при резком (десятикратном) снижении численности обыкновенных полёвокподвижность зверьков возросла в три раза. Более высокая подвижность при низкой плотности была зарегистрирована и у американских «белоногих мышей». Наблюдения в естественных популяциях полевых мышей в Польше показали, что количество расселяющихся зверьков, оседающих в данном месте, прямо пропорционально числу оседло живущих здесь особей (Р. Trojan, 1965). Эти наблюдения показывают роль интегративного поведения в формировании естественных внутрипопуляционных группировок.
Реальные пути формирования таких групп прослежены, например, при работе по борьбе с грызунами в природных очагах инфекций. Так, монгольские песчанки, выжившие после обработки ратицидами больших территорий, оставили свои участки и перешли к ненаправленным перемещениям, в процессе которых встречались с другими животными и уже группами и вновь переходили к оседлому образу жизни. Характерно, что в период кочевок между песчанками явно преобладали мирные, контактные формы поведения (Д.П. Орленев, 1981; Д.П. Орленев, С.В. Перепадов, 1981). Увеличение подвижности серых крыс после дератизации животноводческих помещений наблюдал А.Н. Козлов (1980), также связывающий это явление с формированием новых групп. «Тяга к контактам» показана и в экспериментах с грызунами, хотя в целом этот вопрос изучен ещё явно недостаточно. Не исключено, что это свойство неодинаково выражено у представителей разного пола и возраста. В опытах с древесной ящерицейв естественных популяциях удаление самок вызывало резкое повышение подвижности самцов: лишь 5% их осталось на своих участках; в тех же условиях удаление самцов не вызывало увеличения подвижности самок.
В сложившихся популяциях активный поиск информации и контактов составляет одну из характерных сторон исследовательского поведения: «патрулирование» участков, а также спонтанные выходы за их пределы с последующим возвращением характерны для хорошо изученных в этом отношении грызунов, а также других млекопитающих, птиц в репродуктивный период и некоторых других животных.
Интеграция у территориальных животных. В результате стремления к установлению контактов с особями своего вида у оседлых животных с интенсивным типом использования территории полное пространственное разобщение участков обитания встречается не так часто. Такой тип структуры свойствен, например, многим птицам, у которых гнездовые участки формируются только на время высиживания и выкармливания птенцов. У большинства млекопитающих и у многих других животных участки соседних особей в той или иной степени перекрываются. При этом активная охрана участка проявляется лишь в отношении территории, на которую не налегают участки соседей, зоны же совместного использования маркируются всеми животными–соседями, а активно защищаются только от «чужаков» (рис. 10.4). В западной литературе понятие «территория» («territory») часто применяется именно в этом, узком, смысле – для обозначения активно охраняемой части участка обитания.

Области перекрывания участков представляют собой своеобразные «зоны контактов», в которых между соседними животными устанавливаются нейтральные или даже дружелюбные отношения на основе индивидуального опознавания. Так, наблюдения за малыми сусликами показали, что на общей части территории зверьки-соседи мирно кормятся рядом, при опасности прячутся в соседние защитные норы, совместно прогоняют с этой территории «чужаков» (А Н. Солдатова, 1962).
Аналогичные наблюдения имеются и по другим видам. Показано, например, что пойманные в природе полевки-экономки при парном объединении в экспериментальной камере демонстрировали преобладание агрессивных контактов, если это были животные, пойманные в разных местообитаниях, и дружелюбных – если это оказывались зверьки с соседних участков (рис. 10.5).
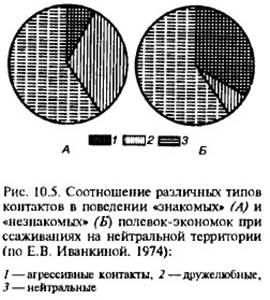
Такие же результаты ранее были получены в опытах с хлопковыми крысами. В экспериментах с разными видами рыб и птиц также было установлено, что предварительное визуальное знакомство снижало агрессивность при последующем ссаживании на общей территории.
Аналогичные зоны перекрывания имеются и у стадных животных. У волков, например, на стыках участков соседних стай имеются особые «буферные зоны», которые посещаются стаями лишь на короткое время и при обилии пищи не используются для охоты. Последнее обстоятельство послужило основанием для предположения о том, что «буферные зоны» функционируют как места беспрепятственного воспроизведения популяций жертв (D. Mech, 1977). У кошачьих лемуров лишь небольшая часть группового участка используется только одним стадом. В местах перекрывания соседние стада контактируют друг с другом, что, в частности, облегчает переход самцов из группы в группу, а соответственно ведет к снижению уровня инбридинга (К. Jones, 1983).
Дифференцированное проявление территориальности с выделением в пределах участка активно охраняемой части открывает возможность «маневра» в использовании пространства: у грызунов, например, при увеличении плотности населения отмечается уменьшение размеров активно охраняемой территории, возрастание степени перекрывания участков и уменьшения их площади (рис. 10.6).

Такая реакция адаптивна на уровне популяции как целого: это предохраняет популяцию от потерь на время, пока включаются и реализуются механизмы приведения плотности населения в соответствии с конкретной экологической обстановкой.
Усилению контактности способствует и различное отношение особей-резидентов к представителям того же вида, проникающим на территорию их участка. Активная охрана территории не абсолютна. Известно, что рецептивные самки бурундуков и бурундуковых белок перестают защищать свои участки от самцов, которых до начала течки на свою территорию не допускают. В большинстве же случаев во все периоды года агрессия по отношению к особям противоположного пола отсутствует или проявляется вяло. У многих видов грызунов участки самок, как и самцов, практически изолированы друг от друга, тогда как участки особей разного пола могут широко перекрываться. Исключение представляют виды, у которых забота о потомстве выражена только у самок: в этом случае в период выкармливания проявляется агрессия самок по отношению к самцам.
У грызунов и некоторых других позвоночных в ряде случаев регистрируется терпимое отношение к молодым животным. Показано, что у грызунов это определяется спецификой их запаха (D. Stoddart, 1974; М. Gоrman, 1982). Этим объясняется слабая выраженность стресса у молодых расселяющихся животных в отличие от взрослых внутрипопуляционных мигрантов, встречающих неизбежную агрессию со стороны резидентов (И.А. Шилов и др., 1969; И.Н. Ивашкина, 1975). Таким путём обеспечивается важная общепопуляционная функция дисперсии молодых особей, способствующей увеличению занятого популяцией пространства.
Внутрипопуляционные группировки. На основе рассмотренных механизмов интеграции общая пространственная структура популяции реализуется в виде системы отдельных внутрипопуляционных группировок, в каждой из которых особи находятся в тесном информационном и функциональном контакте, поддерживая устойчивые взаимоотношения. У высших животных эти отношения строятся на личном опознавании членов группы, и пространственно этологическая структура служит основой стабильности и «управляемости» популяции как биологической системы.
Многочисленные наблюдения за различными видами животных из разных таксонов показывают, что даже в относительно однородных условиях местообитания равномерное или диффузное распределение особей встречается практически как исключение; у самых разных видов явно преобладает агрегированный тип распределения, при котором в составе популяции формируются группировки особей, более тесно связанных друг с другом, чем с другими аналогичными группировками. Их называют демами (J. Gilmur, J. Hes1op-Haккison, 1954), субпопуляциями (С.С. Шварц, 1967), парцеллярными группировками (Н.П. Наумов, 1967, 1971), мерусами (В.Е. Флинт, 1977), внутрипопуляционными группировками (И.А Шилов, 1977). Не вдаваясь в тонкие отличия этих понятий, констатируем лиlliЬ, что внутрипопуляционные группировки представляют собой совокупность особей и семей, живущих в ближайшем соседстве и непосредственно связанных друг с другом теми или иными формами сигнализации. Они возникают активно, и их биологическая роль заключается в различных типах взаимопомощи (Н.П. Наумов, 1967).
Дальнейшие исследования показали, что роль внутрипопуляционных группировок еще более значительна: они представляют собой элементарную структурную единицу популяции, и именно на их основе обеспечивается адаптивный ответ популяции в целом на изменения вне- и внутрипопуляционных условий. Несмотря на индивидуальную неустойчивость самих таких группировок (они могут у ряда видов распадаться и формироваться вновь; состав индивидов в каждой такой группировке может все время меняться), именно их существование обеспечивает устойчивость популяции в целом.
Интеграция особей в составе различного рода внутрипопуляционных группировок всегда сопровождается определённой степенью их пространственного сближения (перекрывание участков, тяга к контактам). Это ведёт к некоторому повышению вероятности обострения внутрипопуляционной конкуренции. Ослабление негативных отношений, повышение рациональности использования ресурсов среды идёт в этом случае через упорядочение взаимных отношений особей в группах. У высших животных это связано с проявлением специфических форм социального поведения, комплекс которого рассматривается как этологическая структура внутрипопуляционных групп.
В общей форме характер взаимоотношений особей во внутрипопуляционных группировках можно определить как более или менее выраженную систему типа «доминирование-подчинение», при которой отдельные особи отличаются в различных видах деятельности. Конкретное выражение форм доминирования-подчинения широко варьирует у разных видов, но всегда основывается на неравнозначности особей, входящих в состав группировки. Такую систему взаимоотношений принято называть иерархической. При такой системе в составе группы выделяются особи разных рангов: «доминанты», «субдоминанты» (подчиняются доминантам, но господствуют над другими) и различные ступени «подчинённых» особей (субординанты).
Такая система иерархии имеет место у многих видов. В частности, она изучена у лабораторных белых мышей. В природных условиях линейный тип иерархии. По-видимому, свойствен серой белке(J. Pack et а)., 1967): у пустынных кавий в Аргентине участки самцов широко перекрываются, и на совместно обитаемой территории среди них устанавливается линейный тип доминирования-подчинения (J. Rood, 1970).
Иерархия такого же типа описана для некоторых рептилий, в частности для ящериц-игуанид, у которых такие отношения формируются в одноразмерных группах особей наряду с территориальностью; доминирующие особи заходят на участки соседей, не подвергаясь нападению с их стороны (D. Hunsaker, В. Burrage, 1969).
Однако в большем числе случаев линейная иерархия замещается групповой, при которой если и выделяется единственный доминант (по крайней мере, в экспериментальных условиях), то остальные особи не составляют линейного ряда соподчинения, а распадаются на две-три группы, в пределах которых животные более или менее равнозначны. Так, в частности, складывается структура отношений у многих видов полёвок и других мелких грызунов.
Есть и иные варианты внутрипопуляционной иерархии. Так, у нильских крокодилов популяция, по-видимому, распадается лишь на две категории особей: доминантные самцы захватывают участки на отмели и защищают их, а все остальные особи участков не имеют. У отдельных видов наблюдается «деспотическое» доминирование одной особи при относительно равнозначном подчинении остальных. Такая структура отношений характерна, например, для хомячков, по крайней мере, в условиях искусственного ограничения территории; близкая к ней – у чёрных крыс. Особенно отчетливо, хотя и в своеобразной форме, такой тип взаимоотношений проявляется у диких домовых мышей, заселяющих замкнутые пространства построек человека.
Отношения доминирования-подчинения чаще наблюдаются только среди самцов, хотя у ряда видов параллельна формируются и среди самок, а у некоторых животных возникает общая система иерархии. Главное биологическое значение структурированности взаимоотношений заключается в том, что при этом повышается степень организованности и «управляемости» группы, что важно при нарастающей по мере совмещения участков вероятности конкуренции. Видимо, поэтому выраженность структуры взаимоотношений тем более отчётлива, чем в большей степени объединяются территории отдельных особей. В частности, деспотическое доминирование одного самца в группах домовых мышей напрямую связано с обитанием в замкнутых ограниченных пространствах: при поселении в природных биотопах этот вид формирует структуру в виде участков отдельных особей с мягкими формами доминирования-подчинения в группах.
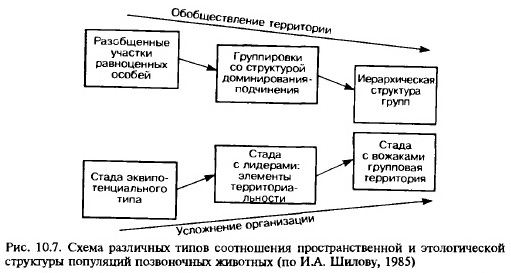
В целом связь системы взаимоотношений в территориальных группах с характером использования пространства можно представить следующей схемой (рис. 10.7):
а) система неперекрывающихся участков обитания. Отчётливо выраженная охрана и маркировка территории; однородность особей по эколого-физиологическим свойствам; отсутствие доминантно-подчинённых отношений между ними (многие коралловые рыбы, неколониальные птицы в период гнездования, млекопитающие семейства куньих и др.);
б) группа перекрывающихся участков обитания. Разделение участков на охраняемую часть и «зоны контактов»; отношения типа доминирования-подчинения, иерархия (многие ящерицы, мелкие грызуны и др.);
в) общая групповая территория. Жесткая иерархия с деспотическим доминированием (домовая мышь в постройках).
Ранговые отличия особей. Иерархическая соподчиненность особей в группах определяет различия в их поведении. Особи-доминанты обладают явными преимуществами и свободно проявляют различные формы деятельности. Так, у мозамбикских тиляпий более крупные самцы, имеющие высокий иерархический статус, чаще занимают территории, имеющие укрытия. В экспериментальных группах грызунов доминанты неограниченно передвигаются по всей территории, беспрепятственно подходят к корму, используют любые убежища и т. д. Подчинённые животные, напротив, ограничены в своих действиях поведением более высокоранговых: их перемещения лимитируются присутствием доминанта, то же относится к использованию укрытий и мест кормления. Чаще всего низкоранговые особи избегают контактов с доминантам, но иногда последний активно сдерживает передвижения подчиненных особей, отгоняет их от кормушек и т. п. При встрече с особями более высокого ранга субординанты демонстрируют особые формы «поведения подчинения», сдерживающие агрессию со стороны доминантов. Вообще, ритуальные формы поведения играют большую роль в снятии агрессии и нормализации деятельности группы в целом.
Нередко в результате таких взаимоотношений у наиболее низкоранговых животных смещается тип суточной активности: в опытных группах такие особи проявляют активность, в том числе кормовую, в периоды, когда неактивны доминантные животные. Нечто подобное можно отметить и в природе: наблюдения за мечеными грызунами показывают, что, в частности, активность молодых самцов, обычно занимающих низшие ранги, не совпадает с активностью взрослых высокоранговых особей либо во времени, либо в пространстве. В некоторых опытах у подчиненных животных отмечалась повышенная исследовательская активность, что можно рассматривать как стимул к миграции. Это подтверждается и наблюдениями в природе: низкоранговые особи, покинувшие свои участки и вошедшие в состав «внутрипопуляционных мигрантов», – обычное явление.
В поведении доминантных особей отмечаются специфические элементы, которые можно рассматривать как проявление некоторых общегрупповых функций. Отмечено, например, что доминирующие самцы домовых мышей активно маркируют и патрулируют территорию группы; то же наблюдается у монгольских песчанок и ряда других видов грызунов и зайцеообразных. У сумчатых летяг самцы-доминанты метят не только территорию, но и подчинённых особей; таким образом индивидуальный запах доминанта приобретает роль фактора групповой интеграции. Подобным же образом поддерживается целостность «кланов» (групп, заселяющих единую систему нор) у барсуков с той разницей, что в этом случае мечение в группе взаимное, без чётко выраженной иерархии.
У некоторых видов существенна роль доминантов во взаимоотношениях с «чужими» особями. Так, у домовых мышей именно доминирующие самцы активно охраняют групповую территорию от вторжения животных извне. Остальные особи, если и вовлекаются в борьбу, то обычно в меньшей степени; при этом активность нападений на «чужаков» обычно коррелирует с положением данной особи в структуре группировки. В семейных группах монгольских песчанок функцию охраны групповой территории выполняют доминантные самки. Аналогичное «разделение обязанностей» в территориальных группах описано и у некоторых других видов.
Как показывают многочисленные наблюдения за искусственными популяциями лабораторных и диких домовых мышей, а также других видов млекопитающих, доминирующие самцы имеют определенное преимущество в половых контактах – как правило, тем большее, чем более четко выражена иерархическая структура. Таким образом, значение внутрипопуляционных группировок как элементарных размножающихся единиц («демов») связано в первую очередь с высокоранговыми самцами.
Иерархическая соподчиненность животных в группах накладывает отпечаток и на их физиологические особенности. В опытах с белыми мышами выяснилось, что у подчинённых особей уровень обмена веществ был ниже, чем у доминантов; при удалении доминантов из групп уровень потребления кислорода у животных, занявших их место, повышался; при рассаживании поодиночке в течение недели повышался уровень обмена и у тех особей, которые не сумели занять высокоранговое положение. Это показывает, что снижение уровня обмена вызвано угнетающим влиянием высокоранговых особей. То же прослежена при исследовании зимних стай синиц-гаичек: у доминантов обмен веществ был выше, чем у других особей; если доминанта удаляли, его место занимала другая особь, после чего уровень потребления кислорода у этой птицы заметно возрастал (О. Hogstad, 1987).
Аналогичные данные получены в опытах с хомяками («оленьи мыши»): уровень потребления кислорода был выше у доминантов, особенно ночью, в период активности. У подчиненных самцов, изъятых из групп, уровень и ритм потребления кислорода постепенно возвращались к норме.
В сложившихся группах различных видов грызунов и некоторых других млекопитающих подчиненные особи всегда проявляют определённо выраженную реакцию стресса, тогда как у доминантов стресс не выражен или выражен слабо. В природных условиях во внутрипопуляционных группировках изученных в этом отношении видов грызунов также отмечается дифференциация особей по уровню стресса, которая в целом соответствует различиям в размерах участков и их расположении. Поэтому есть все основания считать, что и в природе внутрипопуляционные группировки строятся по иерархическому принципу на основе физиологической разнокачественности особей (И.А Шилов, 1972; И.А. Шилов и др., 1973, 1977).
При всем различии особей, составляющих внутрипопуляционную группировку, последняя, несомненно, представляет собой единое целое. Это видно уже из того, что, невзирая на определённую степень взаимного угнетения, отношения в группе более стабильны и терпимы, чем отношение тех же животных к вторгающимся в группу «чужакам». Агрессия по отношению к чужим особям зарегистрирована у многих видов, как в экспериментах, так и в природной обстановке. Показано, в частности, что в природе грызуны, проникающие в состав оседлых группировок, характеризуются повышенным уровнем стресса (И.Н. Ивашкина, 1975). В приближенных к естественным условиям опытах с красно-серой полевкой в Японии было выяснено, что внедряющиеся в сложившиеся группировки новые особи, особенно самцы, занимают в них низшие ранги (М. Kawata, Т. Saitoh, 1988).
Целостность сложившейся группировки прослеживается и в более прямой форме. Так, изучение свободно размножающейся группы кроликов на большом (18 га) выгоне показала, что при нарастании численности популяции, происходящей от трёх исходных пар, практически всё население было сконцентрировано в три группы; обмен особями между ними был ограничен, расселения кроликов по свободной части выгона не наблюдалось (R. Mykitowycz, S. Gambale, 1965). По другим наблюдениям свободно живущая популяция домовых мышей, заселяющая чердачное помещение, распадалась на две группировки, которые не смешивались, несмотря на отсутствие препятствий к передвижению и отдельные переходы зверьков из группы в группу (К. Petrusewicz, R. Andrzejewski, 1962; К. Adamczyk, W. Walkowa, 1971). Подобная же реакция прослежена в эксперименте: при предоставлении двум сложившимся группам домовых мышей доступа друг к другу объединения этих групп не произошло, и особи чужой группы неизменно вызывали агрессивную реакцию (F. Rowe, R. Redfern, 1969). В аналогичных опытах с лабораторными мышами объединение групп состоялось после схватки между доминантами; в образовавшейся таким путём группе все мыши из группировки проигравшего схватку доминанта заняли подчиненное положение (Z. Nowak, 1971).
При контактах стаек чижейчлены стай, постоянно обитающих на данной территории, неизменно доминировали над проникающими в её пределы группами; в то же время внутри стаек сохранялись ранее сложившиеся отношения (J. Senar et al., 1990). Таким образом, в результате тесных взаимодействий в территориальных группировках позвоночных животных формируется определённая разнокачественность особей по физиологическим свойствам, поведению и «вкладу» в общепопуляционные функции. Как будет показано далее, такая функциональная структурированность лежит в основе авторегуляторных реакций на уровне популяции как интегрированной целостной системы.
Груnnировки в nоnуляциях растений. Групповое распределение в пространстве свойственно и структуре ценопопуляций растений. Элементы ценопопуляции размещены в пространстве закономерным образом; распределение субпопуляционных структур связано с поддержанием оптимальной плотности в условиях ценотической конкуренции. Структура популяций растений трёхмерна и может рассматриваться как в горизонтальном, так и в вертикальном векторах. Вертикальная структура связана с размерами и возрастным составом растений; ярусность во многом определяет фотосинтетическую активность ценопопуляции и фитоценоза в целом.
В качестве элементов горизонтальной структуры популяций растений выступают ценопопуляционные локусы (субпопуляции), в роли которых могут рассматриваться любые участки ценопопуляции, отличающиеся по плотности особей, возрастному составу и общей фитомассе.
В пространственной структуре ценопопуляций широкo распространено групповое распределение. Агрегированность растений в популяциях разномасштабна; в популяциях выделяются группы, существенно отличающиеся по размеру. Отличия в размерах группировок и плотности расположения растений в них определяются отчасти неоднородностью условий существования, а отчасти особенностями экологии: дальностью разноса семян и этапом формирования группировки. Последнее отражает динамику структуры, связанную с онтогенезом: развитие скопления начинается, как правило, с образования мелких сгущений за счёт рассеивания семян материнской особи, затем следует развитие молодых растений и появление новых поколений. На этой основе увеличивается плотность и усложняется возрастной состав группировок, которые в конечном итоге смыкаются друг с другом в объединения более высокого уровня. По мере их развития меняется плотность, возрастная структура и степень дискретности растений. Как правило, молодые группировки характеризуются достаточно высокой плотностью и степенью дискретности; старые скопления более изрежены и часто смыкаются с соседними.
Если ценопопуляционные локусы представляют горизонтальную составляющую пространственной структуры, то вертикальная её составляющая представлена ярусным распределением элементов структуры и тесно связана с размерами и возрастным состоянием растений. Так, в лесных фитоценозах, отличающихся наиболее выраженной ярусностью, вертикальная структура отражает распределение крон деревьев разных возрастных групп; в схеме кроны генеративных и сенильных деревьев образуют первый ярус, молодые генеративные – второй, виргильные – третий. Подобная ярусноеть связана с морфофизиологическими отличиями; в частности, разные возрастные группы отличаются строением листьев, что отражает приспособления к различиям условий освещения, влажности и температуры в разных ярусах.
Сходным образом формируется ярусность подземной составляющей пространственной структуры: корневые системы разных возрастных групп «делят» подземное пространство, образуя ярусы, уходящие в глубину почвы.
Структурированность ценопопуляции в пространстве создает условия реализации адаптивных возможностей популяции и устойчивого участия её в биогенном круговороте веществ. На уровне популяции в целом важное значение имеет неоднородностъ возрастного состава отдельных ценопопуляционных локусов, возникающая как результат их асинхронного развития. Возникновение неоднородности основывается на динамике формирования и развития локусов и связано с неравномерностью возобновления на разных участках неодинаковыми темпами развития и отмирания особей в отдельных группировках. Для нормального существования целостной ценопопуляции необходим полный набор разновозрастных локусов; их неоднородностъ и асиихронность развития представляют собой основу устойчивости ценопопуляций (Л.Б. Заугольнова и др., 1988).
Интеграция в стадах в стаях. У номадных групповых животных элементарной единицей популяции является отдельное стадо (стая). Формирование стад как целостных единиц полностью определяется врождённым стремлением находиться в контакте с другими особями своего вида. Это, в частности, выражается в постоянном слежении за соседями, выдерживании внутристадных дистанций и проявлении подражательных реакций, свойственных практически всем стадным животным. На этой основе формируется не только адаптивная пространственная структура стад (стай), но и синхронизация деятельности всех особей, без чего невозможно поддержание целостности подвижной группировки. Степень сложности структуры и внутренней организации стада варьирует у разных видов животных.
Стаи эквипотенциального типа. В относительно просто организованных стаях рыб и некоторых (особенно мелких) птиц каждая особь в равной степени ориентируется на группу ближайших соседей, с которыми находится в постоянном информационном контакте. Особи в составе стаи сходны по своим индивидуальным свойствам и практически равноценны по их экологическому значению для стаи в целом. Такие стаи принято называть эквипотенциальными; особи в них, подражая действиям соседей, одновременно являются объектами подражания для других.
Роль врождённых имитационных рефлексов наглядно продемонстрирована в экспериментах на нескольких видах рыб. Так, в аквариуме, разделённом продольной стеклянной перегородкой, размещались две группы рыб, поддерживающих между собой визуальный контакт. Одна из групп время от времени подвергалась раздражению электрическим током, что вызывало бросок в дальнюю половину аквариума. Вторая, интактная, группа в 83-95 % опытов совершала такой же бросок, имитируя поведение первой. На этой основе оказалось возможным выработать у рыб условный рефлекс, безусловной основой которого является подражательная реакция. Характерно, что такая реакция регистрируется только у стайных видов рыб и проявляется тем более отчётливо, чем более чётко выражена стайность у данного вида.
Отмечается одно важное обстоятельство: подражательная реакция – проявляется сильнее, если объектом для подражания служит группа рыб; на действия одиночной особи такая реакция нередко вообще отсутствовала (Э.Ш. Айрапетянц, В.В. Герасимов, 1965; В.В. Герасимов, 1965, 1967). На этой основе открывается возможность индивидуального маневра отдельных членов стаи в пределах их «жизненного пространства», не нарушающего движения стаи в целом (Е. Shaw, 1965).
У рыб, помимо подражания, определённое значение в интеграции групп имеет химическая коммуникация. На примере золотого карася показано, что если экстракт соскобов эпителия выступает в качестве «вещества испуга», то смывы с поверхности кожи интактной рыбы обладают привлекающим действием и, видимо, способствуют поддержанию целостности стай (Р. Saglio, J. Blanc, 1989).
Многочисленные наблюдения за стаями птиц в природе показывают, что и у них подражательные реакции (часто – в сочетании с активной звуковой сигнализацией) имеют большое значение в интеграции поведения стаи при реакции на опасность, условия кормления, смену направления полета и пр. Эксперименты с большими синицами показали существенную роль взаимного подражания в эффективности питания (J. КrеЬs et al., 1972). Показано также, что объединение птиц в стаи улучшает способность выбора направления полёта при «хоминге» и во время миграций. Причина этого, вероятно, заключается в том, что в стае всегда находятся птицы, быстрее ориентирующиеся и более уверенно выбирающие направление полёта; подражательная реакция, направленная на этих птиц, облегчает ориентацию всей стаи.
При всей однородности составляющих эквипотенциальную стаю особей иногда обнаруживается внутренняя структурированность стаи в виде образования в её составе нескольких группировок. Это характерно как для рыб, так и для птиц. Происхождение и биологическая роль таких группировок не совсем ясны; не исключено, что они имеют определенное гидро- (аэро-) динамическое значение.
Гораздо чаще в таких стаях можно наблюдать «временную разнокачественнос