Часто структурные гены, относящиеся к одному биохимическому пути, объединены в группу, составляющую вместе с оператором единицу транскрипции и регуляции — оперон. Все структурные гены, объединенные в оперон, имеют один операторный участок, локализованной на краю оперона, и координирование регулируются одним репрессором. Оперон представляет собой весьма рациональную и эффективную систему регуляции метаболического пути.
|
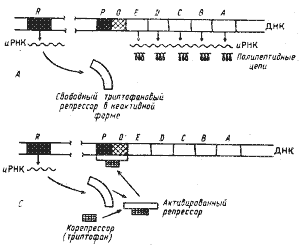
| Рис. 32. Триптофановый оперон Е. coli и механизм репрессии конечным продуктом. А — продукт гена-регулятора (R) — неактивная форма репрессора, неспособная связываться с оператором (О); промоторный участок (Р) открыт: происходит транскрипция структурных генов (Е, D, С, В, А). Б — в присутствии корепрессора образуется активный комплекс корепрессор — репрессор, связывающийся с оператором и закрывающий промотор; транскрипции не происходит (по Lehninger, 1974) |
Процесс транскрипции начинается с прикрепления РНК-полимеразы, катализирующей синтез иРНК, к определенному участку ДНК, называемому промотором (Р). Когда молекула репрессора "садится" на операторный участок, она "закрывает" промотор, тем самым препятствуя связыванию с ним РНК-полимеразы и началу транскрипции. У прокариот пять генов, кодирующих синтез ферментов триптофанового пути, образуют оперон (рис. 32). Ген-регулятор обеспечивает синтез аллостерического белка — триптофанового репрессора, не активного в свободном состоянии. Последний в таком виде не связывается с операторным участком и, следовательно, не может препятствовать началу транскрипции. Когда конечный продукт метаболического пути (триптофан) накапливается выше определенного уровня, он взаимодействует с репрессором и активирует его.
Активированный репрессор присоединяется к операторному участку и подавляет транскрипцию триптофанового оперона. Таким образом, триптофан является корепрессором.
РНК-полимераза – это фермент, который считывает генетическую информацию с ДНК и синтезирует матричную РНК. В РНК-полимеразе есть?-фактор – белок, который распознает промотор и помогает РНК-полимеразе на него сесть и начать транскрипцию. Таким образом переключается работа больших групп генов, это такая системная регуляция. Клетка переключается с одной жизненной программы на другую. Очень большую роль во всех этих процессах играет циклический аденазинмонофосфат (цАМФ). Он – типичный регулятор внутриклеточного метаболизма. Такая система изменения активности аденилатциклазы и, соответственно, концентрации циклического АМФ в клетке, работают не только у бактерий, но и у очень многих организмов, в том числе и у нас с вами. Через аденилатциклазу, регулируя ее активность, действуют некоторые гормоны. Меняя концентрацию циклического АМФ, эти гормоны влияют на внутриклеточные процессы.

Интересно, что циклический АМФ используется не только для регуляции внутриклеточных процессов, но и для межклеточной коммуникации при формировании многоклеточности. Расскажем об этом на примере уникального организма – амебы, которая называется Dictyostelium discoideum. Это одноклеточная амеба, которая живет в почве и питается бактериями. Иногда её относят к грибам.
Принципы регуляции действия генов у эукариот. Регуляторная роль, гистонов, негистоновых белков, гормонов. Особенности организации промоторной области у эукариот.
У эукариот наряду с регуляторными процессами, влияющими на функционирование отдельной клетки, существуют системы регуляции организма как целого. Гормоны образуются в специализированных клетках желез внутренней секреции и с кровью разносятся по всему телу. Но регулируют они процессы синтеза РНК и белков лишь в так называемых клетках-мишенях. Гормоны связываются с белками-рецепторами, расположенными в мембранах таких клеток, и включают системы изменения структуры клеточных белков. Те в свою очередь могут влиять как на синтез белков на рибосомах, так и на транскрипцию определенных генов. Каждый гормон через систему посредников активирует свою группу генов. Так, например, адреналин включает синтез ферментов, расщепляющих гликоген мышц до глюкозы, а другой гормон – инсулин влияет на образование гликогена из глюкозы в печени.
В отличие от прокариот, у которых процессы транскрипции и трансляции не разобщены во времени и в пространстве, у эукариот синтез РНК происходит в ядре клетки, а синтез белков – в цитоплазме. Образующиеся в ядре информационные РНК подвергаются там целому ряду изменений под действием ферментов и в комплексе с различными белками проходят через ядерную оболочку. Разные иРНК транслируются в разное время после их образования. Это зависит от того, с какими белками они связаны в цитоплазме. В отсутствие гормонального сигнала некоторые иРНК остаются нетранслированными долгое время.
Разнообразие форм и функций клеток разных организмов зависит от сложного взаимодействия различных генов между собой и с многочисленными веществами, попадающими в клетку извне или образующимися в ней. Познание регуляторных механизмов транскрипции и трансляции необходимо для управления процессами реализации генетической информации.
С помощью методов молекулярной биологии было исследовано регуляторное действие гистонов и негистоновых хромосомных белков. Как выяснилось, гистоны, особенно гистон H1, оказывают тормозящее действие на ДНК-зависимый синтез РНК. Негистоновым хромосомным белкам тоже приписывают специфические регуляторные функции. Эти белки снимают блокирующее действие гистонов. На их важную роль указывают, помимо прочего, их большое многообразие, неодинаковое содержание их в хроматине различных тканей и на различных стадиях развития, а также результаты экспериментов по реконструкции хроматина. Однако эти данные спорны, так что регуляторное значение гистонов и негистоновых белков остается неясным.
У человека важное значение имеет действие половых гормонов. Развитие первичных мужских половых признаков зависит от образования H-Y-антигена, ген которого, вероятно, находится в половой хромосоме. Стероидные гормоны, вырабатываемые гонадами, транспортируются к клеткам-мишеням, связываются имеющимися там аллостерическими белками- рецепторами, изменяют их конформацию и попадают в виде комплекса гормон-рецептор в клеточное ядро. Происходящая после этого активация транскрипции определенных генов, обусловлена воздействием этого комплекса.
У эукариотической клетки регуляторная область генов устроена гораздо сложнее, чем у прокариот и в ней участвует не только промотор, но и другие элементы которые будут рассмотрены ниже. Но сейчас для демонстрации сложности, о которой идет речь, используем только промотор.
Промотор у каждого из 100000 генов свой, и хотя можно увидеть в разных промоторах общие элементы, и мы с вами их увидим, все эти 100 000 промоторов отличаются друг от друга очень сильно. Их длина варьирует от нескольких сотен до нескольких тысяч пар оснований. На этом пространстве располагаются десятки регуляторных последовательностей различного рода, которые связывают белки, регулирующие транскрипцию. Многие из этих последовательностей определены. Они достаточно коротки и просты. Но если вы обнаруживаете такую последовательность вблизи от какого-то гена, вы ничего не сможете сказать о том, как она влияет на его регуляцию. Требуется экспериментальный анализ, часто очень непростой, чтобы это понять. Потому что различные регуляторные элементы работают сообща, связывают регуляторные факторы, которые взаимодействуют друг с другом, и эти факторы связывают в свою очередь другие факторы, которые также взаимодействуют друг с другом. К тому же это связывание и взаимодействия зависят не только от того, какие именно регуляторные элементы находятся в промоторной области, но и от того, как они расположены относительно друг друга, и от того, какие еще непромоторные элементы, иногда очень удаленные от гена, вовлечены в его регуляцию. См например: Факторы Oct-1 и Oct-2: функционирование в разных типах клеток
Для каждого гена образуется уникальная мозаика регуляторных элементов, действие которых не аддитивно. Одна и таже последовательность, в зависимости от того, какие последовательности находятся вокруг нее, может узнаваться разными белками.
С другой стороны, один и тот же белок может узнавать разные последовательности. Несмотря на интенсивные многолетние исследования функционирования промоторов животных, удалось выработать только самые основные общие представления, которые мало что дают для предсказаний поведения конкретных промоторов. Один и тот же регуляторный элемент может активировать транскрипцию, но может и репрессировать ее, в зависимости от контекста. Загадкой, например, остается почему промоторы определяют однозначное направление транскрипции, хотя многие их составные элементы могут быть направлены по-разному в разных промоторах.
Пример: один из самых известных регуляторных мотивов, так называемый CCAAT бокс [ Mantovani ea 1988 ]. см. Необязательное отступление о происхождении термина бокс
Ну и, наконец, массу проблем вносит то, что транскрипция происходит не просто на ДНК. А на хроматине, который вносит в регуляцию транскрипции еще более чудовищную сложность.