To determine whether there is a certain attribute of hereditary, one can use the method of segregation of genetic markers. It should be established that organisms carrying different alleles of the marker gene also differ in the average values of a certain quantitative trait in the phenotype. If such a pattern takes place, this indicates that the marker gene, which itself does not affect the tag attached to the gene that affects it.
Such a method can be used only in the study of those organisms for which a large number of polymorphic genes are known in different parts of the genome, whose alleles can be determined by a specific method: for example by electrophoretic analysis of protein variants, or in vertebrates by immunological methods. For example, it was shown that in hens with different blood groups the mass of eggs is also different. As far as it is now known, blood group antigens do not affect the size of eggs, it is likely that the genes determining these two traits are closely linked.
Неаллельные гены представляют собой гены, расположенные в разных областях хромосом и кодирующие неравные белки. Неаллельные гены могут также взаимодействовать друг с другом.
В этом случае либо один ген вызывает развитие нескольких признаков, либо, наоборот, одна особенность проявляется под действием набора из нескольких генов.
Существуют три формы и взаимодействия неаллельных генов:
взаимодополняемость;
Эпистаз;
полимерия.
взаимодополняемость
Дополнительное (дополнительное) действие генов является своего рода взаимодействием неаллельных генов, доминантные аллели которых при объединении в генотипе определяют новое фенотипическое проявление признаков. Расщепление F2-гибридов по фенотипу может происходить в отношениях 9: 6: 1, 9: 3: 4, 9: 7, иногда 9: 3: 3: 1.
Примером комплементарности является наследование формы плода тыквы. Наличие доминантных генов А или В в генотипе определяет сферическую форму плода, а рецессивные гены удлиняются. Если присутствует генотип одновременно доминирующих генов А и В, форма плода будет дискообразной. При пересечении чистых линий с разновидностями, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дискообразную форму, а в поколении F2 произойдет фенотипическое расщепление: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 - сферическая и 1 - удлиненная.
эпистаз
Эпистаз - это взаимодействие неаллельных генов, в котором одна из них подавляется другой. Подавляющий ген называется эпистатическим, подавленным - гипостатическим. Если эпистатический ген не имеет своего фенотипического проявления, он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминирующем эпистазе гипостатический ген (B, b) ингибируется доминантным эпистатическим геном (I> B, b). Расщепление фенотипом с доминирующим эпистазом может происходить в соотношении 12: 3: 1, 13: 3, 7: 6: 3. Рецессивным эпистазом является подавление рецессивным аллелем эпистатического гена аллелей ипостатического гена (i > B, b). Расщепление фенотипа может происходить в соотношении 9: 3: 4, 9: 7, 13: 3.
Polymeria - это взаимодействие неаллельных множественных генов, которые однозначно влияют на развитие одного и того же признака; Степень проявления зависит от количества генов. Полимерные гены обозначаются одними и теми же буквами, а аллели одного локуса имеют один и тот же индекс.
полимерия
Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. С кумулятивным (накопительным) полимером степень проявления знака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем более выражен тот или иной знак. Расщепление F2 по фенотипу происходит в соотношении 1: 4: 6: 4: 1.
При некумулятивном полиморфе знак проявляется в присутствии по меньшей мере одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление фенотипом происходит в соотношении 15: 1.
Пример: цвет кожи людей, который зависит от четырех генов.
Наследуемость - это доля изменчивости определенного признака, которая обусловлена наследственными факторами. Понятие наследуемости широко используется в селекции, а также в количественной и популяционной генетике.
Существует несколько типов наследуемости: наследование реализуется - это концепция, используемая селекционерами для оценки целесообразности дальнейшей селекционной работы, она приближена к истинной наследуемости. Истинное наследование делится на два типа: наследуемость в широком смысле и наследуемость в узком смысле (это только та часть изменчивости, которая обусловлена аддитивным действием генов).
Методы оценки наследуемости
Наследуемость можно оценить несколькими способами: во-первых, как описано выше, вы можете рассчитать реализуемую наследуемость; Во-вторых, вклад наследственности и окружающей среды в изменчивость может быть определен путем сведения к минимуму одного из этих факторов; В-третьих, наследуемость может быть исследована путем сравнения родственников и близнецов. В последнее время также используется метод сегрегации маркерных генов.
Минимизация компонентов полной дисперсии
Если один из факторов, влияющих на изменчивость признака, сводится к нулю, то можно определить долю дисперсии из-за другого фактора.
В исследовательских организмах можно установить, является ли определенный признак наследственным по следующему методу: в родительской популяции выберите две группы организмов с экстремальными значениями признака и пересечения между каждой группой. После этого потомство, полученное в результате такой гибридизации, выращивают в тех же условиях. В этом случае экологическая составляющая дисперсии будет равна нулю, и любые различия между двумя классами потомства будут напоминать генетические факторы. Таким образом, если особых различий не наблюдается, это указывает на то, что признак не является наследственным, в противном случае мы можем говорить о наследуемости этого признака.
Аналогичный подход может быть использован для количественной оценки, а именно для определения ценности наследуемости в широком смысле. Генетический компонент изменчивости можно устранить с помощью чистых линий или клонов. Например, Ф. Робертсон изучил изменчивость длины груди у Drosophila: он обнаружил, что дисперсия в генетически гетерогенной популяции составляет 0,366; И в генетически однородных (инбредные линии были использованы) - 0,186. На основании этих данных мы можем вычислить V G:
V G = V Ph - V E = 0,366 - 0,186 = 0,180
Таким образом, наследуемость в широком смысле по длине груди у плодовой мухи такова:
H 2 = 0,180 / 0,366 = 0,49
С другой стороны, наследуемость может быть рассчитана и сведена к минимуму влияние окружающей среды на изменчивость организмов: например, путем выращивания растений в теплицах. Полученные данные сравниваются с дисперсией в популяции, находится в естественных условиях.
Сравнение родственников
Сравнение определенной количественной характеристики у родственников может позволить определить наследуемость в узком смысле. Для этого определяется коэффициент корреляции для этого атрибута (r obs), а также соотношение между родственниками (r exp). Уровень родства отражает пропорцию совместных полиморфных генов у индивидуумов и известен для любой пары родственников (например, для монозиготных близнецов он равен 1 для родителей и их детей, а также для братьев и сестер - 0,5). Отношение этих двух величин дает наследование наследования в узком смысле:
H ^ 2 = frac {r_ {obs}} {r_ {exp}}
Однако сравнение фенотипов родственников, особенно у людей, часто дает очень искаженную информацию о наследуемости, поскольку в семьях, как правило, не только гены, но и окружающая среда являются общими. Таким образом, признаки семьи не обязательно являются наследственными признаками. Иногда их трудно отличить, например, в 1910 году Комиссия здравоохранения США пришла к выводу, что пеллагра является генетическим заболеванием, исходя из того, что на нее повлияли целые семьи. Сейчас известно, что это расстройство связано с дефицитом витамина B 3 и полностью зависит от диеты.
Сравнение родственников также используется для определения наследования личных качеств, таких как фактор интеллекта, склонность к алкоголизму или поведенческие расстройства. Однако наибольшая корреляция между всеми такими характеристиками наблюдается для политических и религиозных взглядов, которые, очевидно, не являются наследственными. Поэтому, чтобы отличить семейные черты от наследственных, для исследования используются часто усыновленные дети и их биологические родители
Сегрегация генетических маркеров
Чтобы определить, существует ли определенный признак наследственности, можно использовать метод сегрегации генетических маркеров. Следует установить, что организмы, несущие разные аллели маркерного гена, также отличаются средними значениями определенного количественного признака фенотипа. Если такая картина имеет место, это указывает на то, что маркерный ген, который сам по себе не влияет на метку, прикрепленную к гену, который воздействует на него.
Такой метод может быть использован только при изучении тех организмов, для которых известно большое количество полиморфных генов в разных частях генома, аллели которых могут быть определены определенным методом: например, путем электрофоретического анализа вариантов белка или У позвоночных иммунологическими методами. Например, было показано, что у кур с разными группами крови масса яиц также различна. Насколько известно теперь, антигены группы крови не влияют на размер яиц, вполне вероятно, что гены, определяющие эти две особенности, тесно связаны.
2.
Polymorphism (multiformity) - any variety of forms of the same species of organisms. Polymorphism is the most universal phenomenon of life. J.B.S. Haldane called man the most polymorphous species on Earth. In humans, almost all signs are polymorphic (eye color, hair, nose and skull shape, blood type, etc.). Polymorphism can be the result of both a discrete intrapopulation variability of a hereditary nature, and can be determined by the rate of the reaction.
Genetic polymorphism arises from the consolidation of different mutations in a population. Therefore, it is classified into: gene, chromosomal and genomic.
Gene polymorphism is due to the presence of two or more alleles. For example, the ability of people to taste the taste of phenylthiourea is determined by a dominant allele (TT, Tt), recessive homozygotes (tt) - it is not felt. Inheritance of blood groups determine three alleles - A, B, O. Chromosomal polymorphism is associated with chromosomal aberrations, and genomic - with changes in sets of chromosomes in a karyotype (heteroploidy).
Polymorphic genetic systems according to their supposed nature include three groups of polymorphisms: transient, neutral, balanced.
Transit polymorphism is explained by a change in the genetic composition of the population at the locus in question. One new allele in the changed environment conditions becomes more profitable and replaces the "original" one. Such polymorphism can not be stable because, due to natural selection, sooner or later the "original" allele will be replaced by a new one and the population will be monomorphic in the "new" allele. The speed of such a process can not be noticed during the lifetime of one generation.
With neutral polymorphism, due to random stochastic processes (gene drift, founder effect), a random change in allele frequencies occurs. For example, the appearance of differences in adaptive-indifferent characters (adherent or loose earlobe). Changes in gene frequencies according to these characteristics are carried out by the mechanism of gene drift, which explains the neutral type of their evolution.
Balanced polymorphism is a polymorphism caused by a complex balance between selection against both homozygotes in favor of heterozygotes. The recessive genotype undergoes a stronger elimination than the dominant one. Differences in the rate of elimination of these two genotypes maintain a constant, stable equilibrium existence in the population of both alleles with the own for each frequency. This explains the stability of such polymorphism. The most fully studied systems of balanced polymorphism associated with selection for malaria - abnormal hemoglobins, thalassemia, deficiency of erythrocyte enzyme glucose-6-phosphate dehydrogenase. The stability of these polymorphisms disappears in connection with the success of the fight against malaria. Balanced polymorphism turns into a transient one. However, to reduce the gene frequencies of already completely pathological genes, since there is no need to protect against malaria, several dozen generations must pass.
A large number of currently open polymorphic systems in humans with a significant number of alleles leads to the fact that almost every person has a unique set of genes, which allows us to talk about the biochemical and immunological personality of the individual. This is of great importance in medical practice, especially in forensic examination.
Полиморфизм (многообразие) - любое разнообразие форм одного и того же вида организмов. Полиморфизм - это самый универсальный феномен жизни. J.B.S. Холдейн назвал человека самым полиморфным видом на Земле. У людей почти все признаки полиморфны (цвет глаз, волосы, нос и форма черепа, тип крови и т. Д.). Полиморфизм может быть результатом как дискретной внутрипопуляционной изменчивости наследственного характера, так и может определяться скоростью реакции.
Генетический полиморфизм возникает в результате консолидации различных мутаций в популяции. Таким образом, он классифицируется на: ген, хромосомный и геномный.
Полиморфизм гена обусловлен наличием двух или более аллелей. Например, способность людей пробовать вкус фенилтиомочевины определяется доминантной аллелью (TT, Tt), рецессивными гомозиготами (tt) - она не ощущается. Наследование групп крови определяет три аллеля - A, B, O. Хромосомный полиморфизм связан с хромосомными аберрациями, а геномный - с изменениями наборов хромосом в кариотипе (гетероплоидии).
Полиморфные генетические системы в соответствии с их предполагаемой природой включают три группы полиморфизмов: переходную, нейтральную, сбалансированную.
Транзитный полиморфизм объясняется изменением генетического состава популяции в рассматриваемом локусе. Одна новая аллель в изменившихся условиях среды становится более прибыльной и заменяет «оригинальную». Такой полиморфизм не может быть стабильным, потому что из-за естественного отбора рано или поздно «оригинальная» аллель будет заменена новой, и популяция будет мономорфной в «новом» аллеле. Скорость такого процесса не может быть замечена в течение жизни одного поколения.
При нейтральном полиморфизме из-за случайных случайных процессов (дрейф генов, эффект основателя) происходит случайное изменение частот аллелей. Например, появление различий в адаптивно-индифферентных персонажах (прилипшие или рыхлые мочки ушей). Изменения в частотах генов в соответствии с этими характеристиками осуществляются механизмом дрейфа генов, который объясняет нейтральный тип их эволюции.
Сбалансированный полиморфизм является полиморфизмом, вызванным сложным балансом между отбором против обоих гомозигот в пользу гетерозигот. Рецессивный генотип подвергается более сильной элиминации, чем доминирующий. Различия в скорости элиминации этих двух генотипов поддерживают постоянное, стабильное равновесное существование в популяции обоих аллелей с собственными для каждой частоты. Этим объясняется стабильность такого полиморфизма. Наиболее полно изучены системы сбалансированного полиморфизма, связанные с селекцией на малярию - аномальные гемоглобины, талассемия, дефицит фермента эритроцитов глюкозо-6-фосфатдегидрогеназой. Стабильность этих полиморфизмов исчезает в связи с успехом борьбы с малярией. Сбалансированный полиморфизм превращается в переходный. Однако для снижения частоты генов уже полностью патологических генов, поскольку нет необходимости защищать от малярии, должно пройти несколько десятков поколений.
Большое количество открытых в настоящее время полиморфных систем у людей со значительным числом аллелей приводит к тому, что почти каждый человек имеет уникальный набор генов, что позволяет нам говорить о биохимической и иммунологической индивидуальности человека. Это имеет большое значение в медицинской практике, особенно при судебной экспертизе.
POPULATION GENETICS
Section of genetics, which studies the gene pool of populations and its change in space and time. Let us examine this definition in more detail. Individuals do not live alone, but form more or less stable groups, jointly mastering the habitat. Such groups, if they are self-reproducing in generations, and not supported only by alien species, are called populations. For example, a herd of salmon spawning in one river forms a population, because the descendants of each fish year after year, as a rule, return to the same river, to the same spawning grounds. In agricultural animals, the population is considered to be a breed: all individuals in it are of the same origin, i.e. Have common ancestors, are kept in similar conditions and are supported by a single selection and breeding work. In aboriginal peoples, the population is members of related camps. In the presence of migrations, the boundaries of populations are blurred and therefore indeterminate. For example, the entire population of Europe - the descendants of the Cro-Magnon people who inhabited our continent tens of thousands of years ago. The isolation between the ancient tribes, intensified with the development of each of them own language and culture, led to differences between them. But their isolation is relative. Constant wars and seizures of territory, and in recent times - huge migration led and lead to a certain genetic rapprochement of peoples. The above examples show that the term "population" should be understood as a grouping of individuals associated with a territorial, historical and reproductive community. Individuals of each population differ from each other, and each of them is something unique. Many of these differences are hereditary, or genetic - they are determined by genes and passed from parents to children. The set of genes in individuals of this population is called its gene pool. In order to solve the problems of ecology, demography, evolution and selection, it is important to know the features of the gene pool, namely: how great is the genetic diversity in each population, what are the genetic differences between geographically separated populations of one species and between different species, as the gene pool is changed by the environment, How it is transformed in the course of evolution, how hereditary diseases spread, how effectively the gene pool of cultivated plants and domestic animals is used. Population genetics is also engaged in studying these issues.
Mutations. When gametes are formed, random events occur - mutations, when the parent allele, say A1, turns into another allele (A2, A3 or any other) that was or did not exist before in the population. For example, if in the nucleotide sequence "... TTST TGG...", encoding the region of the polypeptide chain "... serine tryptophan...", the third nucleotide, T, as a result of the mutation was transmitted to the child as C, then in the corresponding Section of the amino acid chain of the protein synthesized in the child's body, alanine would be located in place of serine, as it is encoded by a triplet of TACs (see HEREDITY). Regularly occurring mutations and formed in a long series of generations of all species inhabiting the Earth, then the huge genetic diversity that we are now witnessing. The probability with which a mutation occurs is called frequency, or rate, of mutation. The rates of mutation of different genes range from 10-4 to 10-7 per generation. At first glance, these values seem insignificant. However, it should be borne in mind that, firstly, the genome contains many genes, and, secondly, that the population can have a significant number. Therefore, some gametes always carry mutant alleles, and in almost every generation there is one or more individuals with mutations. Their fate depends on how much these mutations affect the fitness and fertility. The mutational process leads to an increase in the genetic variability of populations, counteracting the effect of gene drift.
ГЕНЕТИКА НАСЕЛЕНИЯ
Секция генетики, которая изучает генофонд популяций и его изменение в пространстве и времени. Рассмотрим это определение более подробно. Индивиды живут не одни, а образуют более или менее стабильные группы, совместно осваивающие среду обитания. Такие группы, если они самовоспроизводятся в поколениях и не поддерживаются только чужеродными видами, называются популяциями. Например, стадо нереста лосося в одной реке образует популяцию, так как потомки каждой рыбы из года в год, как правило, возвращаются к одной и той же реке на одни и те же нерестилища. В сельскохозяйственных животных популяция считается породой: все индивиды в ней одного происхождения, то есть имеют общих предков, содержатся в одинаковых условиях и поддерживаются одним селекционным и селекционным трудом. В коренных народах население является членами связанных лагерей. При наличии миграций границы популяций размыты и поэтому неопределенны. Например, все население Европы - потомки кроманьонцев, населявших наш континент десятки тысяч лет назад. Изоляция между древними племенами, усиленная развитием каждого из них, своего языка и культуры, привела к различиям между ними. Но их изоляция относительна. Постоянные войны и захват территории, а в последнее время - огромная миграция, привела к определенному генетическому сближению народов. Вышеприведенные примеры показывают, что термин «популяция» следует понимать как группу лиц, связанных с территориальным, историческим и репродуктивным сообществом. Лица каждой популяции отличаются друг от друга, и каждый из них является чем-то уникальным. Многие из этих различий являются наследственными или генетическими - они определяются генами и передаются от родителей к детям. Набор генов у особей этой популяции называется генофондом. Для решения проблем экологии, демографии, эволюции и отбора важно знать особенности генофонда, а именно: насколько велико генетическое разнообразие в каждой популяции, каковы генетические различия между географически разделенными популяциями одного Видов и между различными видами, так как генофонд изменен окружающей средой, как он трансформируется в ходе эволюции, как распространяются наследственные болезни, насколько эффективно используется генофонд культурных растений и домашних животных. В изучении этих вопросов занимается также популяционная генетика.
Мутации. Когда гамет образуется, возникают случайные события - мутации, когда родительский аллель, например A1, превращается в другой аллель (A2, A3 или любой другой), который был или ранее не существовал в популяции. Например, если в нуклеотидной последовательности «... TTST TGG...», кодирующей область полипептидной цепи «... серин триптофан...», третий нуклеотид, Т, в результате мутации был Передается ребенку как C, то в соответствующем Разделе аминокислотной цепи белка, синтезируемого в теле ребенка, аланин будет расположен вместо серина, поскольку он кодируется триплетом TAC (см. HEREDITY). Регулярно происходящие мутации и образуются в длинной серии поколений всех видов, населяющих Землю, а затем огромное генетическое разнообразие, которое мы сейчас наблюдаем. Вероятность, с которой происходит мутация, называется частотой или скоростью мутации. Скорости мутаций разных генов варьируют от 10-4 до 10-7 на поколение. На первый взгляд эти значения кажутся незначительными. Однако следует иметь в виду, что, во-первых, геном содержит много генов, и, во-вторых, что популяция может иметь значительное число. Поэтому некоторые гаметы всегда несут мутантные аллели, и почти у каждого поколения есть один или несколько особей с мутациями. Их судьба зависит от того, насколько эти мутации влияют на физическую форму и плодовитость. Мутационный процесс приводит к увеличению генетической изменчивости популяций, противодействуя эффекту дрейфа генов.
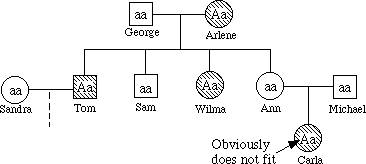
3. Recessive
Fill in the genotypes of the individuals whose genotypes you know. What genotypes are possible for each of the other individuals?
If alkaptonuria is recessive George must be a carrier. See below.

If alkaptonuria is dominant Carla could not have the disease, as indicated in the pedigree chart, since the parents do not express the trait. See Below.