ТЕМА: НАСЛЕДОВАНИЕ КАЧЕСТВЕННЫХ ПРИЗНАКОВ У РЫБ
Основные закономерности наследования генов, расположенных в аутосомах и в половых хромосомах всех диплоидных двуполых организмов полностью приложимы к рыбам.
Наследственная изменчивость у рыб так же многообразна, как и у других животных. Можно выделить четыре группы наследственных различий.
1. Качественные морфоанатомические признаки альтернативного характера, наследуемые в соответствии с законами Менделя и дающие расщепление в потомстве при скрещиваниях особей, различающихся по этим признакам.
2. Количественные различия по многочисленным морфологическим и физиологическим признакам, наследуемые полигенно. Такие признаки зависят в своем выражении не только от многих генов, но и от многих изменчивых факторов внешней среды. Взаимодействия генов, влияющих на один и тот же признак, носят часто аддитивный (суммирующий) характер. Возможны и более сложные неаддитивные взаимодействия: доминирование - подавление одним геном другого, «аллельного» ему гена; эпистаз - подавление геном проявления неаллельного гена; наконец, так называемое сверхдоминирование - усиленное проявление признака у гетерозиготы по сравнению с обеими гомозиготами.
К типичным количественным признакам, варьирующим у большинства видов рыб, относятся: число позвонков и лучей в плавниках, число чешуи и жаберных тычинок, экстерьерные особенности, длина и вес тела, потребление кислорода и устойчивость к высокой и низкой температуре.
3. Биохимические различия, выражающиеся в изменчивости по группам крови и в наличии нескольких форм одного и того же белка, синтезируемых под контролем разных генов или различных аллелей одного гена. Наличие множественных форм белков - изозимов, или изоформ, установлено сейчас практически почти для всех генов, кодирующих ферменты и другие белки Аллельные белковые различия наследуются сходно с различиями по качественным признакам и подчиняются, как и они, менделевским закономерностям.
4. Фенодевианты - уродства и разнообразные отклонения от нормы, обычно плохо проявляющиеся и сложно наследующиеся. Фенодевианты нередко наблюдаются в природных популяциях рыб; еще чаще мы встречаемся с ними в популяциях одомашненных видов. Частота их встречаемости и степень изменения признака в значительной степени зависят от остального генотипа и от среды. Родственное размножение (инбридинг) и неблагоприятные условия существования способствуют появлению фенодевиантов и усиливают выражение признака. Генетический анализ признаков такого типа обычно очень труден.
НАСЛЕДОВАНИЕ КАЧЕСТВЕННЫХ ПРИЗНАКОВ
У ОБЫКНОВЕННОГО КАРПА (CYPRINUS CARPIO L.)
Среди промысловых рыб карп был первым объектом генетических исследований.
Генетика различий в чешуйном покрове. Сплошной чешуйный покров у карпа доминирует над зеркальным типом чешуи. У карпа было выделено две пары аутосомных, не сцепленных друг с другом генов, определяющих характер чешуйного покрова S- и N-. Возможны следующие генотипы и фенотипы карпа (рис. 1):
SSnn, Ssnn.... чешуйчатые,
ssnn………….. разбросанные зеркальные,
SSNn, SsNn ………. линейные зеркальные,
ssNn …………………. голые, или кожистые.


Рис. 1. Типы чешуйного покрова у карпа. а - чешуйчатый (SSnn и Ssnn); б - разбросанный (ssnn); в - линейный (SSNn и SsNn); г - голый, или кожистый (ssNn).
Карпы с генотипами SSNN, SsNN и ssNN нежизнеспособны, эмбрионы, получившие два гена N, погибают на стадии вылупления или вскоре после выхода личинки из оболочки. Результаты скрещиваний:
1) ♀ ssNn х ♂ ssnn → ssnn + ssNn;
голая разбро- разбро- голые
санный санные
2) ♀ ssNn х ♂ ssNn → ssnn + 2 ssNn + ssNN.
голая голый разбро- голые погибают
санные
Повышение температуры во время эмбрионального развития приводит у карпа к выходу эмбрионов из оболочек на более ранних стадиях развития; в результате гибель летальных гомозигот происходит уже после выклева. При пониженных температурах эмбрионы гибнут еще в оболочках.
Ген S доминантен по отношению к гену s; при скрещивании двух гетерозиготных чешуйчатых карпов мы получаем в потомстве классическое менделевское отношение 3: 1, в возвратном скрещивании - 1:1. Результаты трех скрещиваний приведены ниже:
1) Ssnn х Ssnn - 15690 чешуйчатых (75.9%)+4980 разбросанных (24.1%) (Кирпичников, 1948);
2) Ssnn х Ssnn = 3526 чешуйчатых (76.4%)+ 1089 разбросанных (23.6 %) (Probst, 1953);
3) Ssnn X ssnn = 3616 чешуйчатых (50.5%)+3544 разбросанных (49.5%) (Probst, 1953).
Разбросанные карпы (ss) вполне жизнеспособны, но в случае неблагоприятных условий содержания количество чешуйчатых особей повышается при возвратном скрещивании до 52-55 % за счет их более высокой жизнестойкости.
Доминантность гена S является, неполной, так как гетерозиготные чешуйчатые карпы растут несколько быстрее гомозиготных; у гетерозигот чаще наблюдаются нарушения в расположении чешуи на теле.
Различия по жизнеспособности между карпами с генотипами Nn и nn гораздо больше; гетерозиготы отличаются сильно пониженной выживаемостью даже при сравнительно благоприятных условиях выращивания карпов наблюдается довольно существенное отклонение от ожидаемых отношений. Так, при скрещивании дикого сазана с голым карпом можно было ожидать появления чешуйчатых и линейных потомков в равных отношениях.

На самом деле уже в возрасте 3-4 месяцев среди гибридных мальков обычно встречается до 60 % чешуйчатых особей и лишь около 40 % линейных.
Очевидно, гены N и s возникли как две независимые мутации вскоре после одомашнивания европейского сазана Cyprinus carpio carpio. Результатом мутации S→s было появление зеркальных карпов разбросанного типа; мутация n→N привела к возникновению линейных карпов. И разбросанные и линейные карпы характеризуются частичной редукцией количества чешуи и увеличением их размеров. Позднее совмещение двух мутантных генов у одной особи дало голых карпов (ssNn), почти совсем, а иногда и совсем лишенных чешуи.
Разбросанные, линейные и голые особи найдены и в Японии среди карпов, принадлежащих к другому, давно изолированному от европейского подвиду - С. carpio haematopterus. Наследуются эти типы чешуи у европейских и японских карпов одинаково. Описаны разбросанные и линейные карпы и среди вьетнамских карпов С. carpio viridiviolaceus = fossicola, хотя анализ их наследования не произведен.
Таким образом, гомологичные гены чешуи появились у карпов, относящихся к трем подвидам, которые не только имеют разные ареалы, но и очень сильно морфологически и физиологически отличаются друг от друга. Мы видим здесь яркий пример проявления у рыб закона гомологичных рядов наследственной изменчивости, открытого Н. И. Вавиловым (1920).
В таблицах 1 и 2 приведены результаты всех возможных скрещиваний карпов с разным типом чешуйного покрова (в скобках даны теоретические частоты). Расщепление во всех случаях соответствовало ожидаемому.
Таблица 1 – Результаты скрещиваний карпов с разными типами чешуйного покрова
| Родители | Чешуйчатые | Разбросанные | Линейные | Голые |
| ♀ линейная (SsNn) х ♂ голый (ssNn) | ||||
| (725) | (725) | (1450) | (1450) | |
| ♀ линейная (SsNn) х ♂ линейный (SsNn) | ||||
| (2223) | (741) | (4447) | (1482) | |
| ♀ линейная (SsNn) х ♂ линейный (SsNn) | ||||
| (301) | (100) | (602) | (201) |
Таблица 2 - Наследование чешуйного покрова у карпа
| Родители (независимо от пола) | Количество потомков (%) | |||
| чешуйчатые | разбросанные | линейные | голые | |
| Чешуйчатый х Чешуйчатый | ||||
| — | — | |||
| Чешуйчатый х Разбросанный | — | — | — | |
| — | — | |||
| Чешуйчатый х Линейный | — | — | ||
| 37.5 | 12.5 | 37.5 | 12.5 | |
| Чешуйчатый х Голый | — | — | ||
| Разбросанный х Разбросанный | — | — | — | |
| Разбросанный х Линейный | — | — | ||
| Разбросанный х Голый | - | — | ||
| Линейный х Линейный | 33.3 | 66.7 | ||
| 8.3 | 16.7 | |||
| Линейный х Голый | 33.3 | — | 66.7 | — |
| 16.7 | 16.7 | 33.3 | 33.3 | |
| Голый х Голый | — | 33,3 | — | 66,7 |
Различия в выживаемости между карпами, несущими ген и лишенными этого гена, резко усиливаются при неблагоприятных условиях содержания рыб. В опыте, проведенном в 1940 г., потомство от скрещивания чешуйчатого и голого карпов (♀ Ssnn х ♂ ssNn) было посажено в три пруда, сильно различавшихся о своей рыбопродуктивности. Ниже приводится количество выловленных сеголетков (в %).
| Чешуйчатые | Разбросанные | Линейные | Голые | |
| В благоприятных условиях | 27.8 | 25.4 | 24.4 | 22.4 |
| В средних » | 34.8 | 29.4 | 18.4 | 17.4 |
| В плохих » | 38.4 | 36.5 | 14.0 | 11.1 |
Пониженная выживаемость карпов с геном N является следствием неблагоприятного действия этого гена на большое число признаков. Аллели другого гена, S и s, также обладают плейотропным эффектом, но он выражен значительно слабее. Многие органы карпа изменяются под влиянием генов чешуи, меняются при этом и морфологические и физиологические особенности (табл. 3). По весу чешуйчатые карпы обычно оказываются немного лучше разбросанных, в особенности если их не подкармливают. Линейные и голые карпы растут медленнее других, причем это отставание усиливается при недостатке корма. Несмотря на замедленный рост линейных и голых карпов коэффициенты использования корма у них более высокие. Отличаются они и повышенным жировым обменом - летом у линейных и голых карпов жир накапливается быстрее, зимой расходуется в больших количествах, чем у чешуйчатых и разбросанных. С этой особенностью связана, вероятно, и пониженная зимостойкость сеголетков, имеющих ген N.
Таблица 3 – Плейотропный эффект генов чешуи у карпов
| Признаки | Фенотипы и генотипы карпов | |||
| чешуйчатый, SSnn, Ssnn | разбросанный, ssnn | линейный, SSNn, SsNn | голый, ssNn | |
| Вес сеголетков, благоприятные условия * | 93—96 | 85—88 | 79—80 | |
| То же, неблагоприятные условия * | 83—94 | 42—70 | 37—72 | |
| Вес двухлетков * | 94—96 | 86—91 | 83—84 | |
| Среднее число ветвистых лучей в спинном плавнике (D) | 18.8 (17—22) | 18.7 (17—22) | 16.4 (12—19) | 15.4 (5-18) |
| То же, в анальном плавнике (А) | 4.96 | 5.00 | 3.82 | 3.56 |
| Среднее число лучей в брюшном плавнике (V) | 8.91 | 8.68 | 8.76 | 8.47 |
| Среднее число ветвистых лучей в грудном плавнике (Р) | 14.7 | 14.3 | 14.3 | 13.1 |
| Число жаберных тычинок, вариация средних | 24.6—25.1 | 24.3—24.8 | 19.4—21.6 | 18.5—20.5 |
| Среднее число жаберных лепестков | 88.6 | 83.5 | 82.3 | 83.2 |
| Среднее число глоточных зубов | 9.22 | - 9.58 | 7.63 | 7.44 |
| Индекс прогонистости, 1/Н (отношение длины тела к максимальной высоте тела), вариация | 2.33—2.77 | 2.26—2.74 | 2.35—2.86 | 2.35—2.82 |
| Способность к регенерации плавников * | ||||
| Отношение длин задней и передней камер плавательного пузыря | >1 | <1 | ||
| Количество эритроцитов (млн/см3) | 1.93 | 1.99 | 1.76 | 1.69 |
| Гемоглобин (г/%) | 9.02 | 8.87 | 8.18 | 8.28 |
| Устойчивость к нагреву (критическая температура, °С) | 37.6 | 37.5 | 36.8 | 36.6 |
| Устойчивость к дефициту кислорода, выживаемость (мин) | ||||
| Иммунологическая реактивность | Быстрая | Быстрая | Медленная | Медленная |
| Устойчивость к заболеванию краснухой | — | Повышенная | — | Пониженная |
| Интенсивность жирового обмена | Низкая | Низкая | Высокая | Очень высокая |
| Общая жизнеспособность сеголетков, оптимальные условия * | 91—98 | 87—93 | 80—92 | |
| То же, неблагоприятные условия * | 93—95 | 36—37 | 28—60 |
Примечание. * — в % от принятого за 100 значения данного признака у чешуйчатых карпов.
Редукция жаберного аппарата и уменьшение числа глоточных зубов, возможно, являются главными факторами, обусловливающими замедленный рост линейных и голых карпов. Вместо обычных для сазана трехрядных зубов (формула 1.1.3—3.1.1) у карпов этих двух групп часто встречаются двурядные и даже однорядные зубы, зубная формула при этом очень разнообразна (1.3—3.1.1, 1.3—3.1, 1.3—3 и даже иногда 3—3).
Очень характерно действие генов s и N на плавники карпа. У разбросанных карпов число мягких лучей в спинном плавнике оказывается обычно несколько сниженным по сравнению с чешуйчатыми карпами. Уменьшено у них и число лучей в брюшном и грудном плавниках. При наличии гена N действие гена s проявляется сильнее, редукция всех плавников (спинного, анального, брюшных и грудных) у голых карпов (s, N) выражена в значительно большей степени, чем у линейных (S, N). Редуцирующее влияние гена N оказывается, однако, во много раз большим. У чешуйчатых и разбросанных карпов плавники имеют нормальное строение. У линейных и голых карпов некоторые мягкие лучи в средней, а иногда и в задней части спинного плавника не развиваются совсем и общее число лучей заметно уменьшается. Спинной плавник приобретает своеобразную форму (рис. 2). Сильная редукция захватывает обычно и анальный плавник, в меньшей степени затронуты брюшные и грудные плавники. Уменьшено и число жестких лучей в спинном и анальном плавниках.

Рис. 2. Редукция спинного плавника у линейных и голых карпов.
Ген N приводит к нарушению гомеостаза развития у карпа. Это особенно очевидно при изучении взаимодействия генов N и s. На фоне гена N (у гетерозигот Nn) резко усиливается действие гена s, относительно слабо проявляющегося у гомозигот nn. Это усиление действия сказывается на многих признаках, в том числе на количестве жаберных тычинок, содержании эритроцитов в крови и интенсивности жирового обмена.
Представляют большой интерес различия между двумя группами карпов, Nn и nn, по устойчивости к повышенной температуре и дефициту кислорода, по количеству в крови эритроцитов и гемоглобина и по способности плавников к регенерации. Линейные и голые карпы уступают по всем этим показателям чешуйчатым и разбросанным, различия статистически достоверны.
Удивительно широкое поле действия гена N можно объяснить лишь тем, что он работает уже на очень ранних стадиях развития и затрагивает весьма существенные формообразовательные процессы. Действие гена N связано с дефектом в развитии мезенхимы. Ген N представляет собой, очевидно, крупную мутацию, скорее всего хромосомную перестройку типа делеции, затрагивающую небольшой участок хромосомы. У гомозигот NN, вероятно, полностью прекращен синтез одного или нескольких жизненно важных белков, и гомозиготные особи выжить не могут. Сильно нарушен белковый синтез и у гетерозигот.
Хозяйственная ценность разных форм карпа различна. По скорости роста чешуйчатые карпы в большинстве случаев оказываются несколько лучше разбросанных, а линейные - лучше голых, но бывают и обратные соотношения. Вместе с тем линейные и голые карпы всегда растут медленнее чешуйчатых и разбросанных. Отставание карпов с геном N по скорости роста и жизнеспособности усиливается при неблагоприятных условиях выращивания.
Среди многих пород карпа имеются чешуйчатые и разбросанные формы. Встречаются в некоторых породах и голые карпы. Особенно низкой устойчивостью к заболеваниям и к зимовке отличаются линейные карпы. В настоящее время от разведения линейных карпов рыбоводы отказались.
Большая изменчивость мутантных форм карпа сказывается и на самом чешуйном покрове. Количество и расположение чешуи у зеркальных и голых карпов зависит от ряда генов-модификаторов, наследование которых пока детально не прослежено. Крайними вариантами среди разбросанных карпов являются так называемые крупночешуйчатые карпы, с телом, сплошь покрытым чешуей, и рамчатые карпы, имеющие лишь широкую «рамку» чешуи по периферии (рис. 3). Не менее разбросанных изменчивы и линейные карпы (рис. 4).


Рис. 3, 4. 3 - Наследственная вариация в расположении и количестве чешуй у разбросанных карпов (ssnn); 4 - Наследственная вариация в расположении и количестве чешуи у линейных карпов (SSNn и SsNn).
При скрещивании карпа с карасем Carassius carassius и линем Tinea tinea гены чешуи расщепляются по тем же правилам, что и у карпа; эти виды имеют, очевидно, гомологичные гены, расположенные в гомологичных хромосомах. В потомстве от скрещивания карася с голым карпом появляются почти поровну чешуйчатые и линейные гибриды.
Линейные карпокараси очень похожи внешне на линейных карпов (рис. 5), но плавники у них имеют нормальное строение (как и у вьетнамских линейных карпов). Потомки от скрещивания голого карпа с линем также делятся на две группы, но вместо линейных гибридов появляются разбросанные.

Рис. 5. Гибриды карпа Cyprinus carpio с карасем Carassius carassius (по: Lieder, 1957): а — чешуйчатый; б — линейный.
Генетика различий в окраске.
Большое разнообразие окрасок характерно для всех подвидов сазана и породных групп карпа. Генетически хорошо изучены следующие формы.
1. Голубые карпы, часто встречающиеся среди различных одомашненных породных групп. Наследуется голубая окраска как простой рецессив. Немецкие голубые карпы (ген blD) почти не отличаются по жизнеспособности и скорости роста от обычных карпов. Во 2-м поколении наблюдается следующее соотношения фенотипов.
| Поколение | Не голубые, шт. (%) | Голубые, шт. (%) |
| F2 Fb | 4925 (76.0) 757 (50.0) | 1553 (24.0) 756 (50.0) |
В данном случае голубая окраска - результат недоразвития кристаллов гуанина (редукция гуанофоров) в коже карпов - так называемой алампии.
Польские голубые карпы, найденные в прудовом хозяйстве Очаби, очевидно, представляют собою другую мутацию (blp). Это тоже рецессивный ген, но он обладает сильным плейотропным действием. На первом году жизни эти карпы растут быстрее своих нормальных сверстников. Превышение в весе сеголетков голубых карпов можно принять равным 10-20 %. На втором, а в особенности на третьем и четвертом годах жизни рост голубых карпов резко замедляется. Вес трехлеток, в среднем - 1812 г (голубые) и 2687 г (не голубые).
Голубой израильский карп (ген bli) также является рецессивной мутацией. Рост гомозигот по этому гену замедлен, жизнеспособность снижена.
2. «Золотые» особи - точнее, красные или оранжевые, с черными глазами - встречаются во многих странах, как среди культивируемых пород карпа, так и среди разновидностей дикого родича карпа — сазана. Четкое наследование по рецессивному типу характерно для израильских «золотых» карпов (ген g), они лишь немного уступают по темпу роста и жизнеспособности неокрашенным карпам. Ген g также служит для маркирования селекционных линий при промышленном скрещивании.
3. Оранжевые японские карпы — результат взаимодействия двух рецессивных генов b1 и b2 генетическая формула этих карпов — b1b1b2b2. При скрещивании друг с другом гетерозигот по генам b1 и b2 (B1b1B2b2) только шестнадцатая часть потомков получает оранжевую окраску.
Карпы с дупликатными генами b1 и b2 были завезены из Японии в СССР и в некоторые страны Восточной Европы. В Венгрии эти гены получили обозначение p и r. На личиночной стадии двойные гомозиготы по генам b1b2 прозрачны (в коже нет черных пигментных клеток — меланофоров), лишь глаза у них черные (рис. 6). Позднее на теле мальков появляются отдельные пигментированные участки, карпы становятся пятнистыми, оранжево-черными (пегими). Благодаря раннему проявлению и легкой идентификации ген b1 и b2 могут оказаться очень полезными при проведении ряда селекционных экспериментов.
Специализированные породы красных и оранжевых карпов есть на острове Ява в Индонезии (икан-мас и лаок-мас) и в Японии (хигой). Дикие красные карпы найдены в горных районах Вьетнама, в некоторых водоемах их количество доходит до 40-50 %. Единичные экземпляры красных или темно-красных карпов встречаются в немецких прудовых хозяйствах, обнаружены они и в озерах Северной Америки.
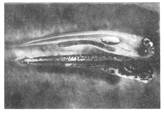
Рис. 6. Прозрачная (генотип b1b1b2b2) и нормальная (В1В2) личинки карпа (фот. В. Я. Катасонова).
Закономерности наследования всех этих «золотых» мутантов не установлены. Можно предположить, что в большинстве случаев мутация связана с блокированием (в большей или меньшей степени) образования меланина и развития меланофоров.
4. Стальная окраска - рецессивная мутация (ген r), обнаруженная в потомстве японских декоративных карпов. У «стальных» карпов уменьшено количество красных и желтых пигментных клеток — ксанто- и эритрофоров. Наследуется ген r очень четко, на жизнеспособность карпов почти не влияет и используется для маркирования селекционных отводок. Сочетание генов r, b1 и b2 дает белых карпов, лишенных и меланофоров, и ксантофоров.
5. Серые и желтые карпы - обычные варианты окраски европейских пород карпа; различие четко выявляется по окраске брюшка. Генетический анализ пока не проведен, но есть основания считать, что различие между ними определяется не более чем двумя-тремя генами. Серая окраска израильских карпов зависит от наличия одного рецессивного гена gr, служащего, как и гены g (золотые) и bl1, (голубые), маркером исходных линий при проведении промышленных скрещиваний.
6. Светлые карпы - доминантная мутация, распространенная среди японских декоративных карпов (рис. 7). Карпы, гомозиготные по этому мутантному гену (LL), погибают на личиночной и мальковой стадиях развития, среди сеголетков их не остается. Гетерозиготы (Ll) выживают, но отличаются пониженной жизнеспособностью.
Ген L в гетерозиготном состоянии оказывает на карпов сильное плейотропное действие — удлиняются грудные плавники, укорачивается задняя камера плавательного пузыря, увеличиваются размеры головы. У светлых карпов удлинен кишечник, в сыворотке крови содержится меньше белка. На первом году жизни эти карпы отличаются ускоренным (примерно на 20%) ростом и более спокойным поведением. Осветление всего тела связано с устойчивой контракцией меланофоров.

Рис. 7. Мутация «светлый карп» (L) (фот. В. Я. Катасонова). Слева — нормальные особи (ll); справа - гетерозиготы (Ll).
7. Второй доминантной мутацией, обнаруженной у японских карпов-хромистов, является своеобразный светло-желтый рисунок — полоса на спине и орнамент на голове (рис. 8). Карпы с таким рисунком, как гомозиготные, так и гетерозиготные (DD и Dd), вполне жизнеспособны. Во 2-м поколении (после скрещивания гетерозигот) наблюдается расщепление по признаку «рисунок-без рисунка» в отношении 3:1.

Рис. 8. Мутация «рисунок» (генотипы DD и Dd) у карпа (по: Катасонов, 1973).
Ген D, как и ген L, плейотропен — у особей с рисунком голова уменьшена, удлинена задняя камера плавательного пузыря, увеличено число позвонков, экстерьер изменен в сторону сазаньего типа. По скорости роста карпы с геном D мало отличаются от контрольных карпов, не имеющих этого гена.
Встречаются и другие вариации окраски как у карпов, так и у сазанов. Совсем не изучены зеленые и фиолетовые карпы, нередко появляющиеся среди амурских сазанов и в породах, созданных с их участием. Не изучены и многие цветные разновидности карпов и сазанов в тропиках — белые, желтые и лимонно-желтые, фиолетовые, коричневые, настоящие альбиносы и другие. Можно лишь предположить, что большинство из этих вариантов окраски определяется одним, иногда двумя или тремя генами, влияющими на развитие пигментных клеток.
До последнего времени не было обнаружено ни одного случая сцепления генов окраски друг с другом и с генами чешуйного покрова S и N, если не считать предполагаемого сцепления дуплицированных локусов b1 и b2. Отсутствие сцепления обусловливает возможность использования многих из этих генов в качестве маркеров селекционных отводок и породных групп карпа.
Наследование других признаков. Перечислим наиболее интересные случаи.
Карликовый карп «Пизарцовичи» был найден в одном из прудовых хозяйств Польши. Судя по расщеплению в F1, это доминантная мутация, очень сильно замедляющая скорость роста рыб. Морфологически карликовые карпы похожи на нормальных рыб, но имеют несколько меньший рот, очень часто у них наблюдаются уродства позвоночника (сращение тел позвонков и др.).
В Китае, Японии, Вьетнаме и Индонезии имеются карликовые разновидности и породы карпа, приспособленные для разведения в мелководных прудах и на рисовых полях. Среди диких сазанов бывшего СССР также обнаружены очень мелкие формы. В Аральском море найден карликовый камышовый сазан, созревающий при весе 200-300 г. В прудах сеголетки этого сазана росли в несколько раз медленнее сеголетков других форм сазана и карпа, т.е. карликовость оказалась наследственной.
Как рецессивная мутация наследуется отсутствие брюшных плавников. В одном из Озер в системе реки Иллинойс (США) обитает изолированная популяция одичавшего карпа, свыше 40 % рыб этой популяции лишены одного или обоих брюшных плавников. Можно предположить наследственный характер этой редукции, однако у карпа плавники нередко редуцируются в результате воздействия на зародыши и на личинки неблагоприятных условий существования. Таким образом, можно говорить только о наследственном предрасположении к нарушению развития.
В Индонезии, одна из пород карпа (кумпаи), имеет, наряду с прогонистой «линеобразной» формой тела, удлиненные плавники. Это изменение, вероятно, связано с простой генной мутацией.
Среди гибридов 2-го поколения между карпом и амурским сазаном были обнаружены несколько десятков рыб, имеющих добавочный преданальный плавник. Эта явно атавистическая мутация была найдена позднее и в 4-м гибридном поколении. Добавочный плавник состоит из двух-четырех крупных жестких лучей (рис. 9). Карпы с добавочным плавником были вполне жизнеспособны и не отставали по скорости роста от своих нормальных братьев и сестер.


Рисунок 9. Мутации «преданальный плавник» у карпа
Мутация «дельфиноподобная голова» была обнаружена в одном из штаммов французского карпа. Скрещивания с неродственными рыбами показали, что мутация доминантна, но характеризуется неполным проявлением; в F1 лишь 62-76 % особей имеют этот признак. Можно, наконец, предположить сравнительно простую генетическую природу укорочения тела (за счет слияния ряда позвонков) у айшгрундской «тарелочной» породы карпа.
В естественных популяциях сазана также довольно часто встречаются многочисленные и разнообразные по характеру аберрации.
Таким образом, известно о наследовании примерно 15-20 генов морфологических признаков у карпа. Среди них особый интерес представляют летальные гены N (редукция чешуи) и L (осветление окраски), убивающие в гомозиготном состоянии своих носителей. Оба гена обладают чрезвычайно широким плейотропным действием и были в свое время отобраны для разведения карповодами-селекционерами. Снижение плодовитости на 25 % благодаря выщеплению гомозигот по леталю не представляет опасности для карпа, способного дать единовременно до миллиона и больше икринок. Сохранение летальных мутаций в племенных отводках карпа легко может быть объяснено наличием у гетерозигот по этим мутациям некоторых преимуществ. Голые карпы (Nn) пользовались успехом у европейских рыбоводов благодаря почти полному отсутствию у них чешуи. Светлые японские декоративные карпы (Ll) привлекали, очевидно, внимание селекционеров своим необычным внешним видом, в особенности при сочетании с другими, рецессивными, генами окраски.