Согласно гипотезе, выдвинутой в 1902 году Бернштейном, потенциал покоя обусловлен тем, что имеет место разная концентрация калия по обе стороны мембраны. То есть существует какой-то механизм активного транспорта калия из клетки. В результате калий перераспределяется между внешней и внутренней сторонами мембраны, и на мембране возникает потенциал, описываемый уравнением Нернста.
Δφ = RT/zF●In([К+о]/ [К+in]) (5).
Но экспериментальные данные показали расхождение с расчетными при малых концентрациях калия вне клетки, соответствующих реальным (единицы миллимоль).
4. Стационарный потенциал.
Согласно этому представлению, возникающий на мембране потенциал является результатом уравновешивающих друг друга потоков ионов в клетку и из клетки. Если брать во внимание три основных иона: калия, натрия и хлора, то ФNa+ + ФK+- ФCl-=0. Поясним примером. Если у нас непроницаемая для ионов мембрана, то по обе ее стороны концентрируются ионы противоположного знака, что минимизирует разность потенциалов на мембране, каков бы ни был ионный состав вне и внутри клетки. Если мембрана полупроницаемая, и какой – то ион может через нее как-то проникать: пассивно или активно, то в результате создавшегося направленного движения зарядов создается разность потенциалов. Согласно уравнению (7) из Л3:
Ф= -cuRT(dc/cdx)-cuqNa(dφ/dx) = - uRT(dc/dx) – cuzF(dφ/dx), где
u= Dβ/RT, D – коэффициент диффузии.
Такая разность потенциалов будет ослабляться по мере того, как будет ослабевать поток. Если он остается постоянным по какой-то причине, как остается постоянным и градиент концентрации ионов, то не меняется и потенциал. Если существуют потоки нескольких ионов, то возникший потенциал будет результатом суммы потенциалов, создаваемых этими потоками. Он, таким образом, зависит от концентраций ионов по обе стороны мембраны, зарядов ионов и проницаемости мембраны.
Такой потенциал, вызванный потоками ионов через мембрану и не изменяющийся во времени из-за того, что потоки друг друга уравновешивают, назвали стационарным. Для него выведено уравнение:
Δφ = RT/zF●In(Рк[К+о]+РNa[Na+o]+ РCl[Cl-in]/ Рк[К+in]+PNa[Na+in]+ РCl[Cl-o]) (6),
где Р-проницаемость для соответствующего иона. P=Dβ/d, где D – коэффициент диффузии, β – коэффициент распределения вещества между водным раствором и мембраной, d – толщина мембраны.
Соглано данным, полученным на гигантском аксоне кальмара, PK:PNa:PCl = 1:0,04:0,045. Таким образом, разность потенциалов на невозбужденной мембране действительно обусловлена, в основном, калием: разностью его концентраций в клетке и вне клетки, и потоком калия через мембрану (таблица 1).
Таблица 1. Равновесные Нернстовские потенциалы для ионов натрия, калия и хлора в состоянии покоя. ∆φ=RT/zF•In [Э out]/[Э in]
| Объект | Равновесный Нернстовский потенциал, мВ | ||
| К+ | Na+ | Cl- | |
| Мышцы лягушки | -98 | +49 | -105 |
| Аксон кальмара | -90 | +46 | -29 |
Эксперименты с радиоактивными изотопами показали, что ионные потоки через мембрану непрерывны, а равновесие имеет динамический характер.
5. Потенциал действия.
При возбуждении нервных клеток, клеток мышц происходит резкое изменение потенциала между внутриклеточной и наружной средой. Потенциал меняет знак и становится положительным. Этот потенциал называют потенциалом действия. Через определенное время потенциал возвращается к исходному значению (рис.3).
Это изменение потенциала распространяется по нервному волокну в обе стороны как затухающее колебание. Было установлено, что электропроводность мембраны резко возрастает при распространении потенциала действия.

Рис.3. Потенциал действия (спайк).
В изучении ПД большой вклад внесли работы на гигантском аксоне кальмара. Этот аксон достаточно крупный, чтобы было возможно введение электрода внутрь.Величина ПД не зависит от величины стимулирующего сигнала и ПД не возникает, если электрический стимул не достигнет некоторого порогового значения. Но величина ПД не зависит от величины возбуждающего сигнала (Рис.4).

Рис.4. Стимуляция потенциала действия.
За счет каких процессов происходит изменение потенциала при прохождении ПД? Эксперименты с радиоактивными изотопами 42К и 24Na показали, что в момент прохождения ПД (Спайка) происходит интенсивный перенос ионов натрия и калия через мембрану.
Положительный потенциал может быть создан только в результате потока ионов натрия (табл.1). Расчеты показали, что при прохождении одного импульса через каждый квадратный микрон поверхности волокна проходит ок. 20000 ионов натрия и калия.
Но проследить за изменением проницаемости мембран в ходе развития ПД сложно технически, поскольку: 1) ПД протекает за 1-2 мс.; 2) при возбуждении мембраны её сопротивление быстро изменяется, что приводит к перераспределению разности потенциалов между элементами электрической цепи, включающей саму мембрану, источник и измерительный прибор. В связи с этим Ходжкин, Хаксли и Катц применили устройство, поддерживающее потенциал на мембране постоянным.
Плотность тока через мембрану измеряли при постоянном мембранном потенциале. За + принимался ток из клетки, за – ток в клетку. После того, как на мембране потенциал был скачком изменен на +56мВ, регистрировался ток внутрь волокна. Его плотность постепенно снижается и меняет направление (суммарный ток). Если в растворе, омывающем волокно, заменить натрий на другой ион, входящий ток исчезал. Регистрировался только выходящий ток, обусловленный выходом калия из клетки (К+) (рис.5).
Разница между суммарным и калиевым током обусловлена входящим током ионов натрия (Na+). Так как потенциал на мембране поддерживался постоянным, то очевидно, что приведённые кривые отражают изменение проницаемости мембраны для ионов (рис.5).
Ходжкин предположил, что под действием деполяризующего потенциала открываются предполагаемые активационные ворота для натрия. Через какое-то время они инактивируются. Наступает рефрактерный период, на время которого генерация очередного ПД невозможна. Это связывают с потерей каналами на какое-то время способности к открытию.

Рис.5. Ионные токи через мембрану при прохождении потенциала действия.
Если в состоянии покоя PK:PNa:PCl = 1:0,04:0,045, то в состоянии возбуждения PK:PNa:PCl = 1:20:0,045.
Удельное сопротивление аксоплазмы равно 10-100 Ом●см. Удельное сопротивление волокна диаметром 1 мкм составляет 109-1010 Ом●см. Удельное сопротивление, в основном, обусловлено липидным бислоем. Но аксон передает импульс на несколько метров без затухания и искажения. Если бы проведение нервного импульса было связано с его передачей вдоль по аксоплазме, то регистрировался бы ток в аксоплазме, а не ионные потоки внутрь и вовне аксона.
Распространению ПД по волокну больше всего соответствует приведенная на рис. 6 электрическая схема-аналог. EK+, ENa+, Ey – “батареи”, обусловливающие движение ионов натрия, калия и других. Сегмент аксона обладает емкостью С, обусловленной липидным бислоем. Падение сопротивления RNa+, обусловливает перераспределение потенциала φ и изменение ионных токов. Таким образом, вдоль аксона запускается процесс изменения потенциала и интенсификации ионных токов наружу и внутрь. Деполяризация определенного сегмента вызывает открытие натриевых каналов и деполяризацию соседних участков.
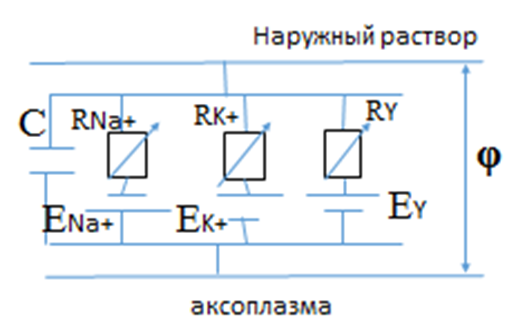
Рис.6. Аналог-схема распространения потенциала действия по нервному волокну.
Для скорейшего распространения импульса по волокну необходимо минимизировать влияние среды, богатой ионами, что способствует снижению Δφ. Это возможно достичь двумя путями: увеличения сопротивления внешней оболочки, либо увеличением диаметра волокна. Второй путь реализуется в аксоне кальмара, имеющего диаметр до нескольких миллиметров.
У высших животных крупные нервы имеют миелиновую оболочку. Она имеет малую емкость. Обмен ионами с окружающей средой обеспечивается только в перехватах Ранвье. Проведение импульса идет путем перескока от одного перехвата к другому.
6. Синаптическая передача.
Когда импульс достигает окончания нервного волокна, он либо переходит на другое волокно, либо на нервно-мышечное соединение. В обоих случаях реализуется синаптическая передача импульса.
Синапс есть функциональный межмембранный контакт двух возбудимых клеток. Расстояние между пресинаптической и постсинаптической мембраной – синаптическая щель, может достигать 10–15 нм. В мионевральном соединении – до 50–100 нм (рис.7).

Рис.7. Синаптическая передача.
Также есть синапсы с сильно сближенными и даже сливающимися пре- и постсинаптическими мембранами. Им соответствуют два типа передачи импульса: химический тип, и прямое электрическое взаимодействие.
Электрическое взаимодействие обеспечивает наиболее быструю передачу импульса, химическое – более избирательную, и возможность более гибкой регуляции постсинаптического потенциала.
Электрические синапсы - это прямые ионпроводящие соединения клеток посредством каналов (коннексонов) в области так называемых щелевых контактов. Они отвечают за проведение импульсов между соседними гладкими или сердечными мышечными волокнами (а кое-где и между нейронами - в сетчатке глаза и в ЦНС), а также обеспечивают коммуникационные функции между соседними эпителиальными или глиальными клетками.
В химических синапсах для передачи информации используются нейромедиаторы (нейротрансмиттеры). Они не только обеспечивают связь в нервной системе, но также служат переключательными элементами. Они могут ускорять или тормозить передачу информации нейроном или передавать ее вместе с другим входящим сигналом.
В химическом синапсе потенциал действия запускает высвобождение медиаторов из пресинаптических терминалей аксона. Затем медиатор диффундирует через узкую синаптическую щель (~ 30 нм), чтобы связаться с рецепторами на пресинаптической мембране нейрона, мышечной или секреторной клетки. В зависимости от типа рецептора и медиатора эффект на постсинаптическую мембрану может быть возбуждающим или тормозным.
Медиаторы высвобождаются путем регулируемого экзоцитоза так называемых синаптических пузырьков. Каждый пузырек содержит некоторое количество («квант») нейромедиатора. В случае двигательной концевой пластинки высвобождается около 7000 молекул ацетилхолина (АХ). Некоторое количество пузырьков, готовых к высвобождению содержимого, скапливается у мембраны в активной зоне. Поступающий потенциал действия служит сигналом для высвобождения нейромедиатора.
Чем выше частота потенциала действия в аксоне, тем больше пузырьков высвобождают свое содержимое (рис.8). Потенциал действия (1) повышает вероятность открывания потенциалзависимых Са2+-каналов в пресинаптической мембране, приводя к увеличению концентрации Са2+ в цитоплазме (2). Внеклеточный Мg2+ ингибирует этот процесс. Са2+ связывается с синаптотагмином (3), который запускает взаимодействие синтаксина и SNAP-25 на пресинаптической мембране с синапто-бревином на мембране пузырьков (4), запуская зкзоцитоз уже заякоренных пузырьков (5) (примерно 100 на один потенциал действия).

Рис. 8. Механизм высвобождения медиатора в синаптическую щель.
Постсинаптическая мембрана содержит рецепторный белок (РБ), пронзающий мембрану насквозь (рис. 9). РБ специфически связывает молекулу медиатора (АХ). В основе связывания диполь-дипольные взаимодействия. В результате РБ изменяет конформацию, открываются ионные каналы (И), что влечет деполяризацию мембраны, генерацию ПД. Молекулы медиатора претерпевают химическую деструкцию под действием специфического фермента (АХЭ).
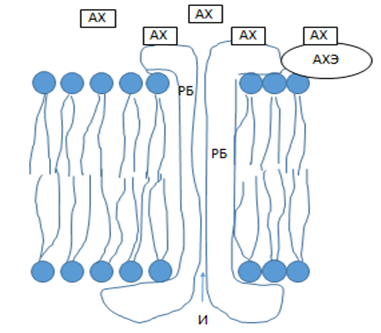
Рис.9. Постсинаптическая мембрана.
Вопросы:
1.Чем обусловлена разность потенциалов между поверхностью мембраны и окружающей средой?
А) Разностью в концентрации ионов по обе стороны мембраны. Б) заряженными группами белков и полисахаридов на поверхности мембраны. В) Заряженными группами белков внутри клетки. Г) Разностью концентраций калия внутри и вне клетки.
2.Что такое донановский потенциал и в каких клетках он наблюдается?
А) Потенциал, устанавливающийся между мертвой клеткой и окружающей средой, при условии, что мембрана проницаема для мелких неорганических ионов, но не проницаема для крупных молекул несущих электрические заряды. Б) Потенциал, устанавливающийся между клеткой и окружающей средой, при условии, что окружающая среда не содержит ионы натрия. В) Потенциал, устанавливающийся вследствие разности концентраций калия внутри и вне клетки. Г) Потенциал между внешней и внутренней средой клетки, возникающий в момент прохождения потенциала действия.
3.Почему поверхностный мембранный потенциал во внутриклеточной и внеклеточной среде убывает с расстоянием в десятки раз интенсивнее, чем внутри мембраны?
А) Из-за большей вязкости мембраны. Б) Из-за меньшей диэлектрической проницаемости мембраны. В) Из-за различного ионного состава по обе стороны мембраны. Г) Из-за того, что концентрация ионов в мембране многократно меньше, чем в омывающей ее среде.
4.Чем обусловлена разность концентраций ионов между внутри- и внеклеточной средой?
А) Наличием внутри клетки белков, имеющих заряженные группы. Б) Действием АТФ-аз и избирательной проницаемостью мембраны для различных ионов. В) Исключительно различной проницаемостью мембраны для различных ионов. Г) Идущими внутри клетки окислительно-восстановительными процессами.
5.Чем индуцируется потенциал действия?
А) Деполяризацией мембраны свыше некоторого порогового уровня. Б) Нарушением действия АТФ-аз. В) Нарушение проницаемости мембраны для хлорида и прочих анионов. Г) Открытием калиевых каналов.
6. Чем обусловлено изменение потенциала на мембране клетки с отрицательного на положительный в момент прохождения спайка?
А) Потоком ионов хлора из мембраны во внешнюю среду. Б) Потоком ионов натрия внутрь клетки. В) Потоком ионов калия из клетки. Г) Изменением структуры мембранных белков.
7.Почему потенциал действия распространяется по нервному волокну?
А) Из-за тока, возникающего в аксоплазме. Б) Из-за последовательной деполяризации соседних участков, что сопряжено с открытием натриевых каналов. В) Из-за тока, возникающего между деполяризованной мембраной и окружающей средой. Г) Из-за тока, возникающего между деполяризованным участком мембраны и аксоплазмой.
8. Что обеспечивает синапс?
А) Межмембранный контакт двух возбудимых клеток. Б) Передачу энергии посредством индуктивного резонанса. В) Ионный транспорт между двумя клетками. Г) Активный транспорт.
9.Как высвобождается нейромедиатор из синаптических пузырьков?
А) В результате пробоя пузырьков под действием разности потенциалов между пре- и постсинаптической мембранами. Б) В результате активного транспорта в синаптическую щель. В) В результате каскада процессов, индуцируемых потенциалзависимым выходом кальция в цитоплазму. Г) В результате эндоцитоза, происходящего на постсинаптической мембране.
10. Какую функцию в нервном волокне выполняет миелиновая оболочка?
А) Ускоряет прохождение нервного импульса. Б) Предохраняет волокно от механических повреждений. В) Обеспечивает питание волокна. Г) Предохраняет волокно от чрезмерной деполяризации.
Глава 5. БИОЭНЕРГЕТИКА