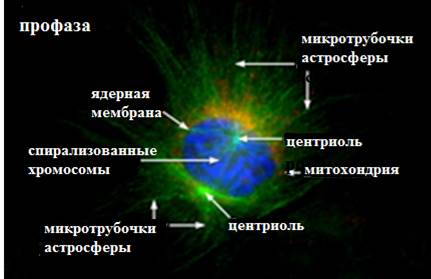
Рис.7. Микрофотография электронограммы профазы митоза, сделанные флюоресцентным микроскопом:
А – прометафаза, Б – метафаза (Из:Интернет-ресурсов с сайта Moleсular Expres- sions /https://micro.magnet.fsu edu/ cells/ fluorescence mitosis/index. html.
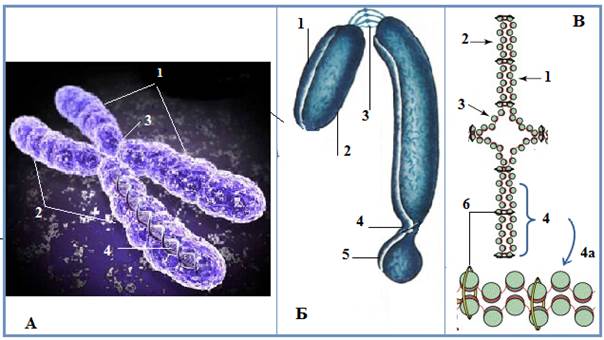 Рис. 8. А – фото метафазной хромосомы (флюоресцирующий микроскоп):
Рис. 8. А – фото метафазной хромосомы (флюоресцирующий микроскоп):
1-2 – левая и правая хроматиды, 3 – центромера, 4 – нить ДНК;
Б – схема метафазной хромосомы: 1-2 - левая и правая хроматиды, 3 – центромера, 4 – вторичная перетяжка, 5 – спутник;
 В – схема соединения двух хроматид в районе центромеры: 1-2 - левая и правая хроматиды, 3 – центромера,; 4 - участок соединения двух хроматид белком когезином(6), тоже в увеличенном виде ((Из: Интернет-ресурсов с сайтов schools.keldysh.ru JPG 400×363; bionet.nsc.ru JPG 455×613; pisum.bionet.nsc.ru JPG 600×485
В – схема соединения двух хроматид в районе центромеры: 1-2 - левая и правая хроматиды, 3 – центромера,; 4 - участок соединения двух хроматид белком когезином(6), тоже в увеличенном виде ((Из: Интернет-ресурсов с сайтов schools.keldysh.ru JPG 400×363; bionet.nsc.ru JPG 455×613; pisum.bionet.nsc.ru JPG 600×485
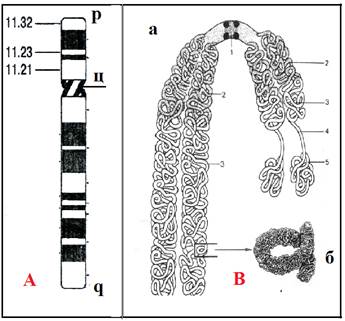
Рис.9. А - схема хромосомы человека (р-короткое и q-длинное плечи, ц – центромера; цифры – номера сегментов и районов плеча) (Из: Клаг,2009);
В - схема метафазной хромосомы: а) 1 - центромера с кинетохором; 2 – гетерохрома - тин; 3 –эухроматин; 4 – вторич- ная перетяжка; 5 – спутник; б) участок с более сильным увеличением (сверхспирализа- ция) (Из: Хадорн, 1988)
Центромера – это место соединения двух хроматид, специфическая область эукариотической хромосомы, в которой хроматин конденсирован в большей степени (рис.9А). В метафазной хромосоме центромера состоит из двух участков хроматид, соединенных специфическими белками (рис.10). Локализация центромеры на метафазной хромосоме определяется первичной структурой ДНК. Центромера содержит специфические тандемные повторы (α-сателлиты ДНК). Их количество и структура и расположение на хромосоме различаются не только у разных видов, но даже у разных хромосом одного вида, определяя тип хромосомы (метацентрические, субметацентрические и акроцентрические).
Центромера играет фундаментальную роль в движении хромосом к полюсам деления и точном распределении дочерних хроматид по дочерним клеткам в процессе митоза и мейоза. В митозе и во втором деле-
нии мейоза цетромеры сохраняют тесную связь между дочерними хромосомами и разделяются только в анафазе. К центромере прикрепляются микротрубочки веретена деления, которые обеспечат расхождения дочерних хроматид к полюсам. Прикрепление микротрубочек осуществляется в особом участке центромеры – кинетохоре, которыйконтактирует с центромерными районами и формируется на период деления клетки (рис.10В-11).

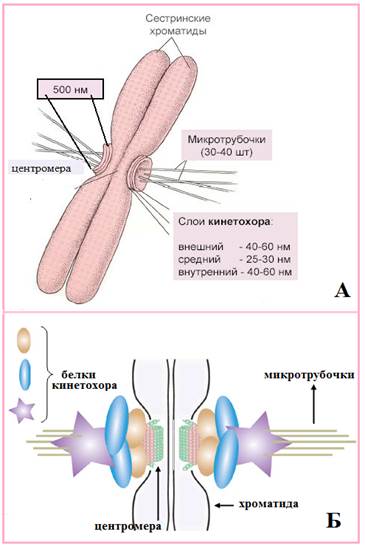
Рис. 10. Центромера и кинетохор: А – электронная микрофотография участка метафазной хромосомы в районе центромеры; Б – схема молекулярного строения центромерных участков двух хроматид, соединения их с кинетохором и микротрубочками веретена деления; В – схема соединения кинетохора с микротрубочкой (Из: Интернет-ресурсов с сайтов Developental Biology and genetics mglinets.narod.ru. JRG 450x507, nanonewsnet.ru JPG 498x313, pisum.bionet.nsc.ru JPG 600x485)
Кинетохор (рис.10Б-11) - это комплекс, состоящий из участка ДНК в районе центромеры, определённых ДНК-связывающих белков, РНК и белка тубулина. Кинетохор имеет вид трехслойной пластины, состоящей из внутреннего (40-60 нм), светло-окрашенного среднего (25-30 нм) и внешнего (40-60 нм) слоев, различающиеся белковым составом (рис.) Метафазная хромосома несет два кинетохора: по одному на каждую из своих хроматид. Кинетохоры связываются с растущими от центриолей на полюсах деления микротрубочками: кинетохор одной дочерней хроматиды связывается с микротрубочками одного полюса, другой дочерней хроматиды - другого полюса (рис.10Б). К каждому кинетохору прикрепляется несколько десятков микротрубочек (рис.11), при этом центромера с одной стороны метафазной хромосомы (одной хроматиды) оказывается связанной с микротрубочками, идущими только от одного из полюсов веретена деления. Контакт кинетохора с микротрубочками стабилизирует микротрубочку, таким образом, что связанные с хромосомами микротрубочки не деполимеризируются.
Таким образом, кинетохор служит:
1 - центром прикрепления микротрубочек веретена деления;
 2 - обеспечивает выравнивание хромосом метафазной пластинки на экваторе деления клетки в метафазе;
2 - обеспечивает выравнивание хромосом метафазной пластинки на экваторе деления клетки в метафазе;
3 - участвует в процессе расхождения хромосом во время анафазы.
Рис.11. Морфологическая(А) и молекулярная (Б) структуры кинетохора. Объяснение в тексте.
(Из: Интернет-ресурсов с сайтов pisum.bionet. nsc.ru JPG 600×485; Barbara Mellone, Sylvia Erhardt & Gary H. Karpen, Nature Cell Biology (2006) nature.com JPG 580×618)
II. Исчезают ядрышки вследствие прекращения синтеза р-РНК и инактивации рибосомных генов.
III. Ядерная ламина (белок ядерного матрикса), связанная с ядерной мембраной разрушается в результате деполимеризации составляющих ее промежуточных филаментов, а ядерная мембрана распадается на мелкие фрагменты и становится невидимой; ядерные поры растворяются в цитоплазме и распадаются на субъединицы (рис.).
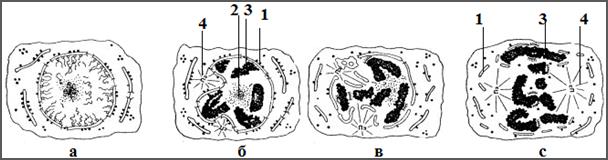 Рис. 12. Изменение состояния ядерной оболочки в процессе интерфазы и профазы митоза: 1 –ядерная мембрана, 2 – ядрышко, 3 – хроматин, 4 - центриоль (Из: Зацепин и др., 1976)
Рис. 12. Изменение состояния ядерной оболочки в процессе интерфазы и профазы митоза: 1 –ядерная мембрана, 2 – ядрышко, 3 – хроматин, 4 - центриоль (Из: Зацепин и др., 1976)
IV. ЭПС и аппарат Гольджи распадаются на везикулы.
V. Центриоли клеточного центра расходятся к полюсам и начина - ют участвовать в формировании веретена деления. Веретено деления образуется из микротрубочек: центром образование становятся центриоли. Микротрубочки веретена деления прикрепляются к центромерам хромосом в области кинетохора (рис.). В дальнейшем кинетохоры сами становятся центрами организации микротрубочек. Центриоли с астросферами, состоящими из астральным микротрубочек и веретена деления образуют митотический аппарат. Длительность профазы – от 2 до 270 м (рис.).
Метафаза
В метафазе все хромосомы выстраиваются на экваторе клетки (рис.6,13). Этому предшествуют события прометафазы: кинетохоры связываются с растущими микротрубочками обеих полюсов и скользят по ним: кинетохор одной хроматиды связывается с микротрубочками веретена деления одного полюса, кинетохор другой хроматиды связывается с веретеном деления другого полюса (рис.14Б,В). Прочное связывание и выравнивание хромосом требует времени. Выравнивание хромосом в метафазной пластинке (т.е. на экваторе) свидетельствует о начале метафазы и необходимо для правильного распределения генетического материала по дочерним клеткам. Хромосомы выстраиваются в экваториальной плоскости, образуя метафазную пластинку хромосом или материнскую звезду.
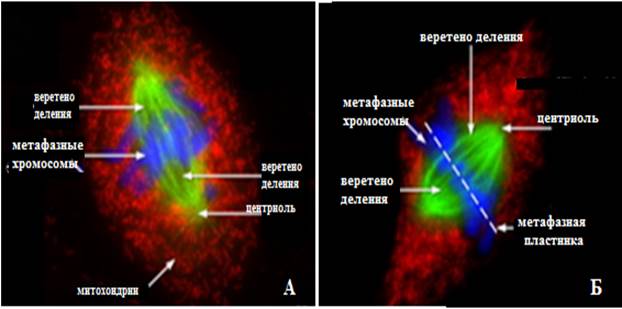
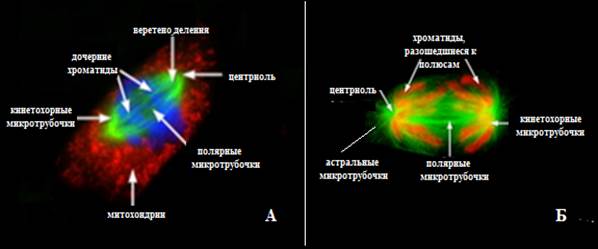
Рис.13. Микрофотография электронограммы метафазы митоза, сделанные при помощи флюоресцентного микроскопа: А – прометафаза, Б – метафаза (Из: Интернет-ресурсы с сайта Molecular Expressions™)
Метафазная пластинка – паспорт организма; число и форма хромосом являются характеристикой вида. Постепенно белок когезин между сестринскими хроматидами разрушается и их разделение завершается к концу метафазы; кажущаяся связь сохраняется (видна только в районе центромеры). Веретено деления полностью сформировано, но остается структурно динамичным: в нем постоянно происходят процессы сборки и разрушения микротрубочек. В метафазе веретено деления состоит из микротрубочек 3 видов: кинетохорных (связывают кинетохор хроматиды с диплосомой), полярных (идут от диплосомы к центру клетки), астральных (направлены от диплосомы к поверхности клетки) (рис.14Б).
Движение кинетохоров по микротрубочкам осуществляют так называемые молекулярные двигатели – ферментные комплексы, которые способны использовать энергию гидролиза АТФ для продвижения по поверхности микротрубочек. Существует два типа двигателей: динеин - крупный ферментный комплекс, который перемещается по микротрубочке к ее минус – концу и кинезин, перемещающийся к плюс-концу. Оба двигателя прочно прикрепляются к хромосомам и тянут их по микротрубочке с большой скоростью. Длительность метафазы – 0,3-175 мин.
Если на этой стадии клеточного деления воздействовать на клетки определенными алкалоидами (колхицин), то дальнейшее расхождение хромосом прекращается и они остаются в зоне экватора в виде метафазной пластинки, при помощи которой изучают кариотип клетки.
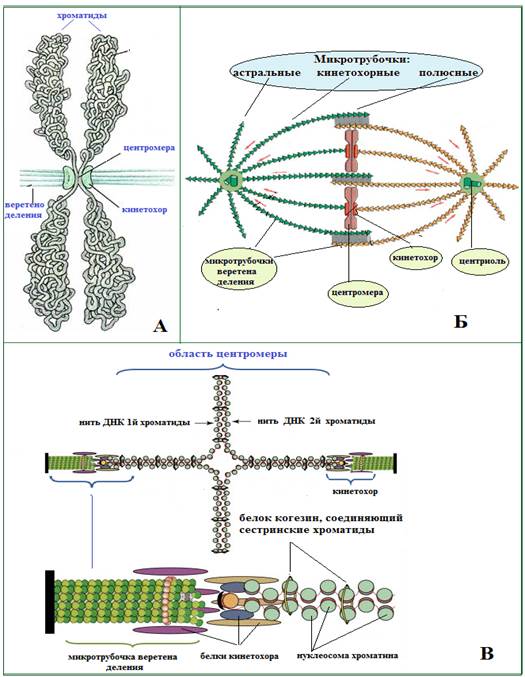
Рис. 14. А – схема присоединения веретена деления к метафазной хромосоме; Б - схема расположения митотического аппарата в районе центромеры метафазной хромосоме; В – схема молекулярной структуры района «центромера-кинетохор-микротрубочка» (Из: Интернет-ресурсы с сайтов: salem.k12.va.us JPEG 300×437 и pisum.bionet.nsc.ru JPG 600×485 )
Анафаза
Хроматиды отделяются друг от друга в области центромеры и начинают расходится к полюсам клетки со скоростью до 1 мкм/мин, при этом центромерные участки обращены к полюсам, а теломерные – к экватору клетки (рис.6,15,16).
Движение хромосом обеспечивается:
1 - изменением длины микротрубочек веретена деления: укорочением кинетохорных и удлинением полярных;
2 - действием молекулярных двигателей, перемещающих хромосомы вдоль кинетохорных и полярных микротрубочки в сторону друг от друга.
 Сигналом к движению является резкое повышение в гиалоплазме концентрации ионов Са2+. Разошедшиеся к полюсам сестринские хроматиды формируют дочерние звезды. Продолжительность – от 0,3 а до 100мин.
Сигналом к движению является резкое повышение в гиалоплазме концентрации ионов Са2+. Разошедшиеся к полюсам сестринские хроматиды формируют дочерние звезды. Продолжительность – от 0,3 а до 100мин.
Рис. 15. Фотографии электронограммы анафазы митоза в животной клетке: А –ранняя анафаза, Б –поздняя анафаза (Из: Интернет-ресурсы с сайта Molecular Expressions™)

Рис. 16. Анафаза в клетках кореш- ка лука: а – ранняя анафаза(1-рас- ходящиеся хроматиды, 2 – стенка растительной клетки); б – поздняя анафаза (3 – дочерняя звезда хрома- тид, 4 – веретено деления) (Из:)
Телофаза
Дочерние звезды формируется вокруг своей центриоли (рис.17). Фрагменты ядерных мембран собираются вокруг каждой хромосомы и сливаются, образуя пузырьки. Субъединицы ядерных пор встраиваются в их стенки. Через поры в пузырьки проникают белки, формирующие промежуточные филаменты, которые образуют ядерную ламину. Таким образом, образуется двойная оболочка вокруг каждой хромосомы. Такая структура называется кариомера (или мини-ядро).
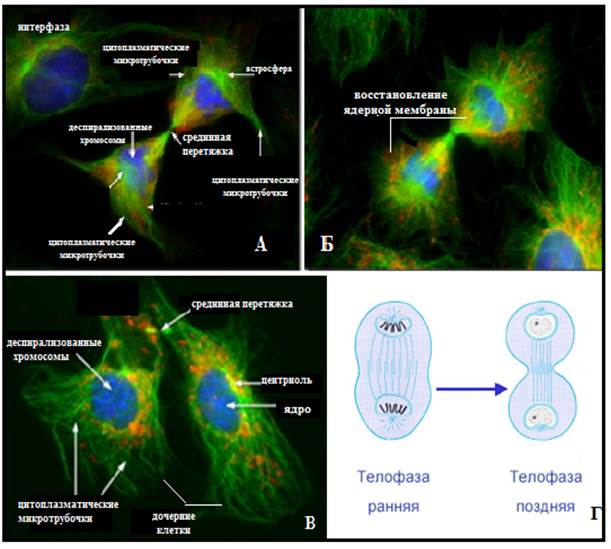
Рис.17. А-В - фотографии электронограммы телофазы митоза в животной клетке: А –ранняя телофаза, Б –поздняя телофаза; В – цитокинез в поздней телофазе; в синий цвет окрашены хромосомы, в красный – ядерная мембрана, в зеленный микротрубочки; Г – схематическое изображение телофазы; (Из: Интернет-ресурсы с сайта Molecular Expressions™)
Затем кариомеры, локализованные вокруг каждой центриоли, сливаются, образуя два дочерних ядра. Хромосомы деспирализуются; формируются ядрышки.
В поздней телофазе завершается цитокинез (цитотомия) - разделение клетки на две дочерние с помощью сократительногоактин–миозинового кольца. Актин-миозиновое сократительное кольцо начи- нает формироваться в поздней анафазе под внутренней поверхностью плазмолеммы в районе экватора. Направление первичной борозды и сократительного кольца определяет расположение веретена деления. Первой на поверхности клетки возникает борозда (рис.18), под ней формируется так называемое сократимое кольцо. Сокращение кольца происходит за счет взаимодействия актина микрофиламентов с миозином, подобно тому как это происходит при мышечном сокращении. Таким образом, деление клетки и механизм движения имеют одинаковую химическую природу.
В результате сокращения кольцо сжимается и стягивает плазмолемму. Это происходит до тех пор, пока мембраны не сольются в центре клетки, разделив цитоплазму и образуя две дочерние клетки. После цитокинеза восстанавливается аппарат Гольджи и ЭПС.

Рис.18. Цитокинез животной клетки: образование актин-миозинового кольца при дроблении бластомеров у лягушки (Из:Интернет-ресурсов с сайта pisum. bionet.nsc.ru JPG 600×485,)
На рисунках 19-20 представлены фотографии процесса митоза и микрофотографии микропрепарата: «Митоз в корешке лука». У животных разделение происходит за счет образования перетяжки между двумя клетками. По мере сокращения кольца клетка разделяется перетяжкой на две, которые в конце концов разделяются, вдобавок оставляя еще небольшое остаточное тельце – связанные друг с другом фрагменты встречных микротрубочек веретена, располагавшиеся первоначально в экваториальной плоскости.
В случае, если в процессе митоза будут иметь место отклонения, то наблюдается явление эндомитоза. Так, в случае разрушения веретена деления, нарушается процесс расхождения хромосом, что приводит к полиплоидии – увеличению количество хромосом, чаще всего кратное двум. Если не происходит цитокинез, то возникают многоядерные клетки (синцитий), которые в дальнейшем могут разделится путем цитокинеза с образованием одноядерных клеток (шизогония у простейших). Эндомитоз приводит к увеличению размеров клетки и ее функциональных возможностей. Вследствие этого его можно рассматривать как механизм приспособления (адаптации) клетки к изменяющиеся условиям среды.


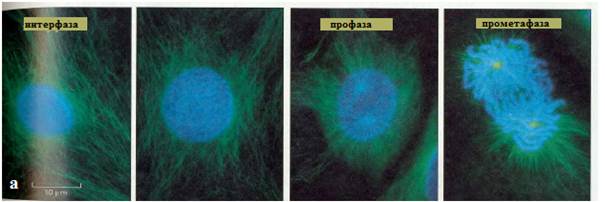 Рис. 19. а – б: микрофотографии процесса митоза, сделанные с помощью флюо- ресцентного микроскопа: синим цветом обозначен хроматин и хромосомы, зеленым – белок тубулин микротрубочек веретена деления (Из: Интернет-ресурсы - фото с сайта Molecular Expressions™)
Рис. 19. а – б: микрофотографии процесса митоза, сделанные с помощью флюо- ресцентного микроскопа: синим цветом обозначен хроматин и хромосомы, зеленым – белок тубулин микротрубочек веретена деления (Из: Интернет-ресурсы - фото с сайта Molecular Expressions™)
Рис.20. Митоз в клетках корешка лука: 1 – интерфаза, 2 – профаза, 3 – поздняя метафаза, 4 – ранняя метафаза, 5 – ранняя анафаза (Из: Интернет-ресурсы с сайта shvedun.ru JPG 1140×760 )
Глава 3. Мейоз
У половых клеток клеточный цикл включает мейоз, особый тип деления клеток, состоящий из двух последовательных делений; при этом репликация ДНК происходит только перед первым – редукционным делением (рис.21-22,таблица 1), включающим сложную профазуI (рис.23). Профаза I состоит из 6 фаз: лептотены, зиготены, пахитены, диплотены, диакинеза. 6 фаза - диктиотена характерна только для оогенеза. На этой стадии хромосомы приняв форму «ламповых щеток» (рис.24), прекращают изменения до наступления у женщин репродуктивного возраста.

Рис.21. Схема мейоза. Красная стрелка показывает начало первого деления, синяя – второго

Рис. 22. Схема процесса мейоза: 1- профаза I (1-центриоль, 2 – ядерная мембрана, 5 – конъюгирующие гомологичные хромосомы на стадии кроссинговера); 2- мета –фаза I (3-веретено деления, 4-бивалент хромосом); 3- анафаза I; 4-телофаза I; 5-профаза II; 6-метафазаII; 7-анафазаII; 8-телофазаII (Из: Интернет-ресурсов с сайта dic.academic.ru JPG 398×399
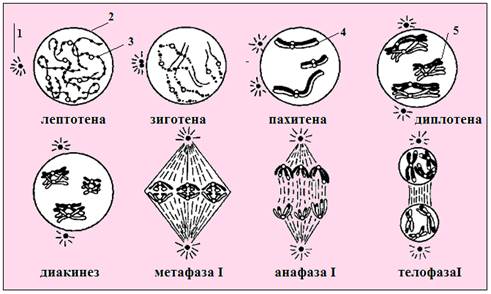
Рис.23. ПрофазаI первого деления мейоза:1 – центриоль, 2 – ядерная мембрана, 3 - хроматин, завершивший спирализацию,4 - гомологичные хромосомы образующие бивалент, 5 – бивалент начавший расхождение (Из: Интернет-ресурсы с сайта bse.sci-lib.com JPG 285×400)

Рис.24. Схема строение бивалента типа «ламповых щеток» (По Callan, 1941; из: Жимулев, 2007).
Содержание событий профазыI мейоза представлено в таблице 1. В профазеI мейоза: образуются биваленты, состоящие из 2 гомологичных хромосом, каждая из которых состоит из двух сестринских хроматид, между которыми происходит кроссинговер – обмен наследственной информацией между гомологичными хромосомами (рис.25). В первом делении мейоза в анафазеI к полюсам отходят гомологичные хромосомы, состоящие из 2 хроматид. Во втором делении мейоза к полюсам расходятся сестринские хроматиды. В результате образуются 4 гаплоидные клетки (рис.26).
В результате мейоза образуются половые клетки с гомологичным набором хромосом, то есть, мейоз приводит к редукции хромосомного набора клетки. После слияния сперматозоида и яйцеклетки в процессе оплодотворения у зиготы восстанавливается диплоидный набор хромосом. Кроме того, в результате кроссинговера генетический материал хромосом обновляется. Кроссинговер приводитк возникновению генетической комбинативной изменчивости и перетасовке отцовских и материнских генов.
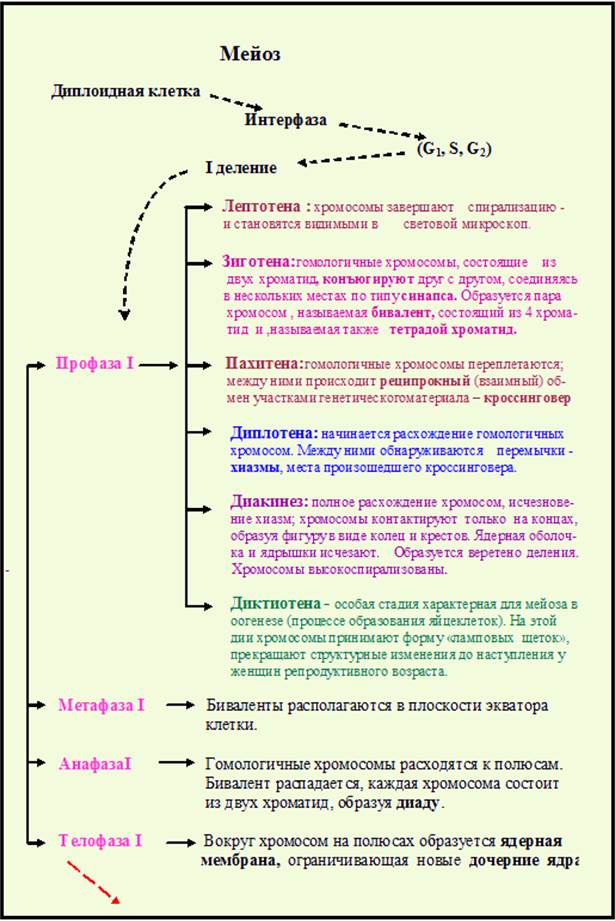 Таблица 1
Таблица 1
Продолжение таблицы 1
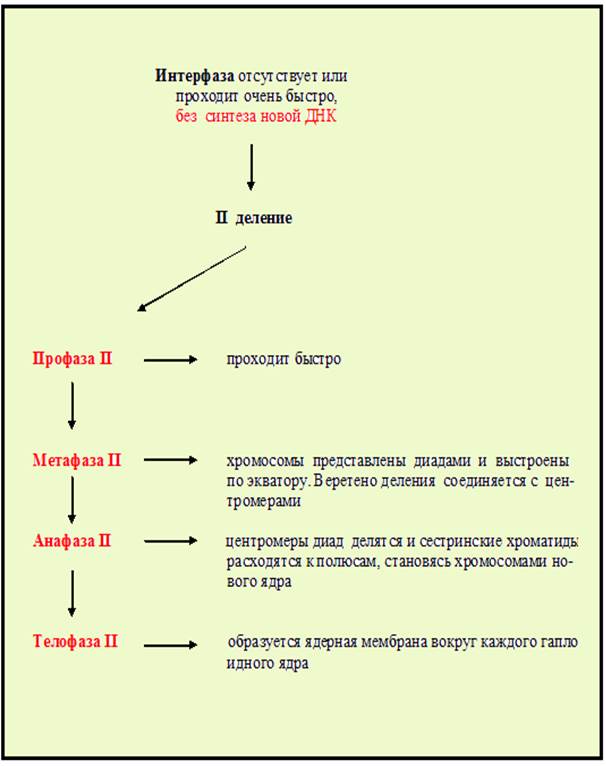

Рис.25. Схема кроссинговера и его результат (Из: Интернет-ресурсов с сайта
evolbiol.ru JPG 720×540,)

Рис.26. Последовательность расхождения хроматид в про - цессе мейоза: бивалент гомоло- гичных хромосом после крос - синговера в метафазеI; расхож- дение гомологичных хромосом; расхождение хроматид в анафа- зеII (Из: Интернет-ресурсов с сайта evolbiol.ru JPG 720×540,)