Транслоказы – это специальные мембранные белки-переносчики, облегчающие диффузию веществ через мембрану. Перенос веществ обеспечивается за счёт временного связывания транслоказы с диффундирующим веществом и не требуют энергии. В своей структуре эти белки имеют особые участки, определенным образом ориентированные на наружную или внутреннюю поверхность мембраны. В отличии от канальных белков транслоказы в ходе переноса веществ претерпевают изменения своей конформации. Это происходит в процессе взаимодействия транслоказы с переносимым веществом (лигандом). Поскольку для транспорта каждой отдельной молекулы или иона переносчик должен изменять свою конформацию, то скорость транспорта веществ с помощью транслоказы в несколько раз меньше, чем это происходит при переносе через каналы.
Транспорт с помощью переносчиков-транслоказ может быть активным и пассивным. В случае пассивного транспорта перенос веществ идет по направлению концентрационного градиента (то есть, из области с большей концентрации в область с меньшей концентрацией) и не требует дополнительных затрат энергии. Транслоказы значительно ускоряют скорость простой диффузии, поэтому такой транспорт называют облегченной диффузией.
Благодаря переносчикам-транслоказам облегченная диффузия идет с большей скоростью, чем обычная диффузия. Транслоказы работают в обоих направлениях в зависимости от концентрации переносимого вещества. Они могут переносить одно вещество в одном направлении (пассивный унипорт, рис.57), два вещества в одном направлении (пассивный симпорт) или два вещества в разных направлениях (пассивный антипорт) (рис.58).
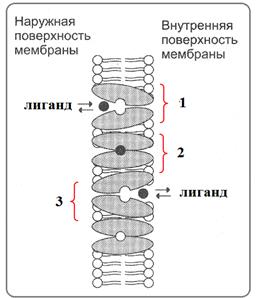 Транслоказы работают следующим образом (рис.57): лиганд (вещество, переносимое транслоказой) взаимодействует с транслоказой на поверхности мембраны или вблизи нее. Затем комплекс транслоказы с лигандом, будучи подвижным в самой мембране, поворачивается на 180˚ к ее противоположной стороне, затем комплекс распадается и высвобождает лиганд во внутреннюю среду. Транслоказа, освободив- шаяся от лиганда, снова переворачивается к внешней стороне мембраны, на которой может соединиться с другим лигандом. Транспорт с участием переносчиков обладает свойством насыщения. Суть этого свойства заключается в том, что при увеличении концентрации лигандов в окружающем растворе скорость их переноса сначала возрастает, а затем остается постоянной. Это объясняется ограниченным количеством транслоказ.
Транслоказы работают следующим образом (рис.57): лиганд (вещество, переносимое транслоказой) взаимодействует с транслоказой на поверхности мембраны или вблизи нее. Затем комплекс транслоказы с лигандом, будучи подвижным в самой мембране, поворачивается на 180˚ к ее противоположной стороне, затем комплекс распадается и высвобождает лиганд во внутреннюю среду. Транслоказа, освободив- шаяся от лиганда, снова переворачивается к внешней стороне мембраны, на которой может соединиться с другим лигандом. Транспорт с участием переносчиков обладает свойством насыщения. Суть этого свойства заключается в том, что при увеличении концентрации лигандов в окружающем растворе скорость их переноса сначала возрастает, а затем остается постоянной. Это объясняется ограниченным количеством транслоказ.
Рис.57.Пассивный унипорт глюкозы(лиганд) в эритроциты с помощью транслоказы ГЛЮТ-1: 1,2 – транслоказа взаимодействует с лигандом; 3 – транслоказа поворачивается на 180˚ и высвобождает лиганд в полость клетки (Из:Северин,2007)

Рис.58. Транспорт анионов НСО-3 и Сl- через мембрану эритроцитов: А - в венозных капиллярах анион НСО3-, образованный при диссоциации Н2СО3, выходит из эритроцита в кровь. В обмен на анион НСО3- транслоказа переносит в эритроцит ион Сl-, Б – в легких обмен ионов идет в обратном направлении. Таким образом, обеспечивает удаление СО2 из организма (Из: Северцев,2007)
Переносчики участвуют в переносе только определенных веществ и, тем самым, обеспечивают избирательность поступления. В тоже время
один и тот же переносчик может обеспечивать транспорт нескольких лигандов. Например, переносчик ионов К+, также способен переносить ионы Na+, но не транспортирует Сl- или незаряженные молекулы сахарозы. Транслоказа, специфичная для нейтральных кислот, перено- сит аминокислоты глицин, валин, но не может транспортировать аспарагин или лизин. Благодаря разнообразию и специфичности транслоказ осуществляется избирательный перенос веществ через мембрану.
Примером транслоказ являются транспортные белки семейства ГЛЮТ. Они обеспечивают облегчённую диффузию глюкозы через мембраны. Существует 5 разновидностей белков ГЛЮТ, обнаруженных во всех видах тканей:
ГЛЮТ-1 (эритроцитарный тип) обеспечивает транспорт глюкозы в глиальные клетки мозга;
ГЛЮТ-2 (печёночный тип) участвует в транспорте глюкозы в кровь из энтероцитов и печени; β-клетки поджелудочной железы;
ГЛЮТ-3 (мозговой тип) обеспечивает приток глюкозы к клеткам нервной и других тканей;
ГЛЮТ-4 (мышечно-жировой тип) является главным переносчиком глюкозы в клетки мышц и жировой ткани и единственным, регулируемым инсулином. Поэтому мышцы и жировую ткань называют инсулинзависимыми тканями.
ГЛЮТ-5 (кишечный тип) обеспечивает транспорт глюкозы в клетках тонкого кишечника.
Все 5 типов ГЛЮТ имеют сходную первичную структуру и доменную организацию. У большинства мембранных белков ГЛЮТ последовательность аминокислот изучена хорошо, а трехмерная структура известна только для некоторых из них (рис.59). Здесь приведена структура переносчика глюкозы, содержащего 12 трансмембранных α-спиральных фрагментов и один олигосахарид, ориентированный в окружающую среду.
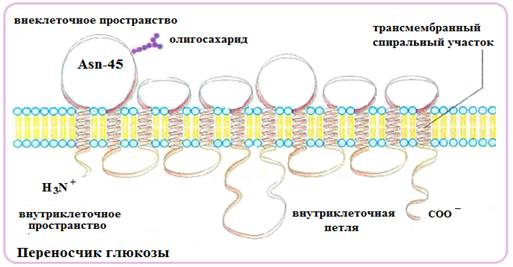
Рис.59. Транслоказа из семейства транспортных белков ГЛЮТ
(Из: Интернет-ресурсов с сайта)
 Для того, чтобы попасть в клетку глюкоза сначала должна связаться со своим переносчиком (ГЛЮТ), локализованным в клеточной мембране. Затем, изменение конформации молекулы переносчика обеспечит перенос глюкозы через мембрану
Для того, чтобы попасть в клетку глюкоза сначала должна связаться со своим переносчиком (ГЛЮТ), локализованным в клеточной мембране. Затем, изменение конформации молекулы переносчика обеспечит перенос глюкозы через мембрану
С помощью транслоказ переносятся вещества не способные к простой диффузии. Исключением является вода, которая переносятся путем простой диффузии, но для интенсификации транспорта воды (например, в почечных канальцах и секреторных эпителиальных клетках) для увеличения скорости её диффузии используется специальная транслоказа - белок аквапорин. Он образует в мембране клетки не- ионные каналы, специальные водные каналы, пропускающие в клетку воду (рис.60).
Рис.60. Водный канал, образованный белком аквапорином по П.Эрг (Из: Интернет-ресурсов с сайта https://kine-ziolog.bodhy.ru, Сазонов, 2011)
Водный канал для проведения только воды, а не ионов и представляет собой цилиндр, по которому движутся молекулы воды. Аминокислоты аквапорина расположены таким образом, что электростатическое поле, создаваемое ими, имеет полярность, которая переключается в центре канала на обратную. Поэтому молекулы воды, дойдя до середины канала, переворачиваются, таким образом, что их дипольные моменты в верхней и нижней части канала оказываются направленными в противоположные стороны. Это переориентирование позволяет предотвращать прохождение через канал заряженных ионов. Такой канал обеспечивает огромную пропускную способность: до миллиарда молекул воды в секунду. В настоящий момент известно уже около 200 разновидностей белков водных каналов у живых организмов и около 11 - у человека. Водные каналы клетки регулируют свой объём и внутреннее давление, а также всасывание воды в почках животных и другие функции.
Итак, транслоказы – это интегральные мембранные белки, имеющие специфичность в отношении переносимых веществ. Они обеспечивают транспорт органических веществ неэлектролитов и ионов, выполняя при этом функцию ферментов, катализирующих транспорт через мембраны. Такой перенос не требует дополнительных затрат энергии, осуществляется по градиенту концентрации в процессе облегченной диффузии. Направление диффузии при этом может менятся в зависимости от изменения градиента концентраций.
Ионные насосы
Наряду с каналами пассивного транспорта веществ, существует каналы активного транспорта, которые называются ионными насосами. Их также называют ионными помпами, ионными обменниками или транспортёрами. Активный транспорт ионов обеспечивает биоэнерге - тику клетки, процессы клеточного возбуждения, всасывания, а также выведения веществ из клетки и организма в целом.
И онные насосы – это крупные трансмембранные ферментные белки, встроенные в мембраны клетки. Они способны связывать и расщеплять АТФ и получившие в связи с этим название АТФазы (аденозинтрифосфатазы). В процессе расщепления АТФ высвобождается химическая энергия, заключённая в них. Эту освобождённую энергию транспортные мембранные АТФазы тратят на перенос определённого вещества против градиента концентрации. Таким образом, они выполняют функцию переносчиков веществ и являются молекулярными транспортёрами веществ через мембрану. Перенос веществ с помощью ионных насосов против градиента концентрации с использованием энергии АТФ называется активным транспортом.
Ионные насосы переносят дискретные частицы - ионы в три стадии: 1 – захват частицы с одной стороны мембраны; 2 - перенос ее через мембрану (транслокация); 3 – высвобождение с другой стороны мембраны. На осуществление этого и расходуется энергия АТФ.
Работа транспортных мембранных АТФаз включает следующие этапы:
1 - фермент АТФаза связывает молекулу АТФ и ионы на поверхности мембраны в центрах связывания АТФ и ионов, расположенных на её поверхности. Ещё один активный центр на поверхности молекулы АТФазы не участвует в процессе гидролиза АТФ и переноса ионов, но имеет значение для регуляции активности фермента. После связывания ионов структура этого активного центра изменяется. Это изменение активизирует АТФазу и запускает процесс гидролиза одной молекулы АТФ. Гидролиз АТФ необходим для использования энергии, заключенной в АТФ и каталитируется АТФазами. Реакция проходит по уравнению:
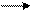 АТФ + Н2О АДФ + Фн (неорганический фосфат) +Q
АТФ + Н2О АДФ + Фн (неорганический фосфат) +Q
2 - гидролиз АТФ осуществляется АТФазой поэтапно: вначале происходит связывание АТФ, фосфорилирование белка и отщепление АДФ, гидролитическое расщепление белок-фосфатной связи и высвобождение неорганического фосфата:
ионы
Е + АТФ → Е-Фн + АДФ,
Е- Фн + Н2О → Е + Фн , где Е - АТФаза, Е-Фн -
фосфорилированный белок
Фосфорилируется карбоксильная группа одного из аминокислотных остатков. Связь фосфата с аминокислотным остатком в фосфори -лированном белке богата энергией и именно эта энергия тратится на активный перенос ионов кальция.
3 - перенос ионов через мембрану. АТФазы в нефосфорилирован- ном состоянии доступны для ионов только с внешней стороны мембраны и недоступны с внутренней. После их фосфорилирования ион-связывающие центры становятся доступными с внутренней стороны и недоступными с внешней. Следовательно, фосфорилирование приводит к переносу центров связывания ионов через мембрану (транслокации) и перемещению белковой молекулы в мембране, что обеспечивается изменением её конформации. Освобождаемая при распаде АТФ энергия и используется для изменения конформации АТФазы. В результате изменения конформации участок фермента, связывающий ион, поворачивается и оказывается по другую сторону мембраны.
5 - изменение прочности связи ионов с ферментом и их высвобождение;
6 - возврат системы в исходное состояние.
В состав АТФаз входят ферментный центр, ионный канал и структурные элементы, которые препятствуют потере ионов во время работы ионных насосов.
Ионные насосы по источнику энергии для своей работы можно разделить на "симпортные" и "антипортные". Симпортные и онные насосы транспортируют в одном направлении два вещества: одно из них имеет большую потенциальную энергию для движения через мембрану, которая используется для транспорта другого вещества. Например, симпорт в клетку глюкозы с помощью ионов натрия, или симпорт ионов кальция с помощью ионов натрия. Антипортные и онные насосы или ионные обменники используют встречный транспорт двух веществ с разной потенциальной энергией диффузии. Так работает, например, натрий-калиевый ионный насос.
Ионные насосы - АТФазы являются одними из самых главных транспортных структур мембраны. Они схожи между собой по строению и механизму действия, но отличаются по специализации, т.е. каждый их вид перетаскивает через мембрану разные ионы. В настоящее время хорошо изучены следующие АТФазы: Na+/K+-АТФаза; Ca2+-АТФаза; H+-АТФаза; H+,K+-АТФаза; Mg2+-АТФаза, которые обеспечивают переме - щение ионов Na+, K+, Ca2+, H+, Mg2+ изолированно или сопряжено. Например, Na+ сопряжённо с К+; Н+ сопряжённо с К+.
- Н+-АТФаза (протонный насос или протонная АТФаза) - это внутриклеточные АТФазы, работающие как протонные насосы: они перекачивают через мембраны ионы водорода (рис.61). В митохондриях в ходе перемещения электронов по дыхательной цепи выделяется энергия, которая переносит протоны из матрикса в межмембранное пространство через протонные насосы. В результате создается протонный градиент (разность концентраций протонов – разность рН и разность электрических потенциалов), энергия которого используется для синтеза АТФ в ходе обратного перемещения протонов в матрикс через другую транспортную систему, так называемую, Н+-АТФ-синтазу. (см.подробнее: «Митохондрии»).
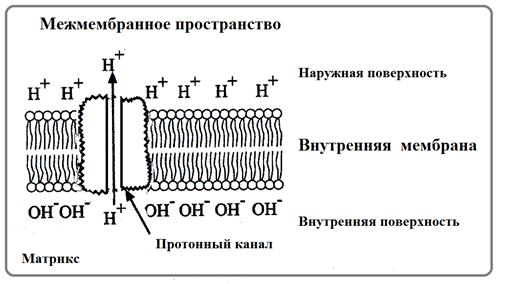
Рис. 61. Н+-АТФаза во внутренней мембране митохондрии (Из: Сеитов,2007)
Встречается в мембранах митохондрий, хлоропластов и бактериальных клеток.
- Н+,К+-АТФаза – ( протонно-калиевая АТФаза ) обеспечивают секрецию соляной кислоты клетками желудка, перемещая на наружную сторону мембраны ионы водорода, которые создают кислую среду в желудке. Н,К-АТФаза работает по принципу обменника, меняя внутриклеточные ионы водорода на внеклеточные ионы калия.
- Са+-АТФаза (кальциевая АТФаза) встречается у мембран мышечных клеток и эритроцитов, а также в мембране эндоплазматической сети и переносит два иона Са2+ из цитоплазмы в пузырьки саркоплазматического ретикулума и внеклеточное пространство (рис.62)
Са+-АТФаза представляет собой группу кальциевых АТФаз, которые различаются по локализации в клетке, строению и способу регуляции. Входя в клетку, ионы Са2+ активируют различные внутриклеточные процессы. Различают Са2+-насосы плазмолеммы клетки и мембраны ЭПС: кальциевый насос плазмолеммы переносит ионы Са2+ из клетки в межклеточное пространство, а цитоплазматический насос - из гиалоплазмы в полость ЭПС, создавая внутриклеточные депо Са2+. В ЭПС мышечных клеток Са2+-АТФаза составляет больше половины белков мембраны и является частью механизма, регулирующего цикл сокращения–расслабления мышечного волокна. Сокращение мышц обеспечивается выходом ионов кальция из саркоплазматического ретикулума и его взаимодействием с сократительными белками актином и миозином. Удаление Ca2+ из цитоплазмы и накопление его в полостях эндоплазматического ретикулума приводят к расслаблению мышц.
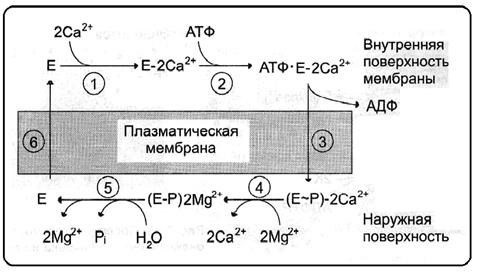
Рис.62. Последовательность работы Са2+-АТФаза:1 – связывание ионов Са2+; 2 – гидролиз АТФ, фосфорилирование и изменение конформации фермента Са2+- АТФазы; 3 – открытие Са2+- АТФазы с наружной стороны и закрытие с внутренней; 4 – высвобождение ионов Са2+ от Са2+ -АТФазы; 5,6 - в присутствии ионов Mg2+ фермент Са2+-АТФаза дефосфорилируется и возвращается в исходное состояние (Из: Северин, 2007)
Концентрация Са2+ во внеклеточной среде и в полостях ЭПС в 10 000 раз превышает концентрацию Са2+ в гиалоплазме, поэтому транспорт ионов Са2+, который обеспечивается работой кальциевых насосов идет против градиента концентрации.
Семейство Са2+ -АТФаз являются трансмембранными белками, состоящими из одной субъединицы, которая пронизывает мембрану несколько раз. Так, например, Са2+-АТФаза саркоплазматического ретикулума поперечнополосатых мышц, пронизывает мембрану a-спиральными участками 11 раз и образует несколько петель, обращенных по обе стороны мембраны (рис.63).
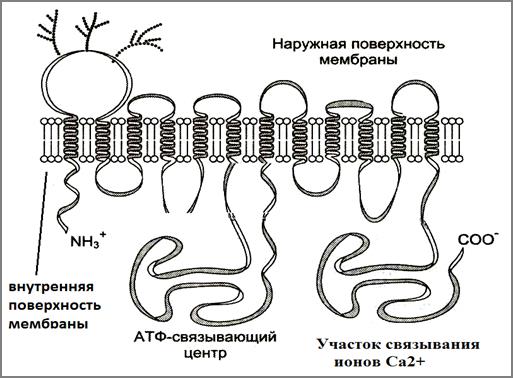
Рис.63.Схема строения Са2+-АТФазы (Из:Северин, 2007)
Две протяженные гидрофильные петли, расположенные со стороны цитоплазмы содержат АТФ-связывающий участок и участок связывания ионов Ca2+. Кроме этих центров, кальциевые АТФазы имеют центр связывания кальмодулина – основного белка, регулирующего плазматическую Са-АТФазу и центр связывания фосфоламбана - белка, регулирующего активность Са-АТФаз мембран ЭПС клеток сердца, печени или эпителия. ЭПС клетки выполняют функцию внутриклеточных депо ионов Ca2+.
Другие Са-АТФазы сохраняют такой же план строения, но отличаются по молекулярной массе.
- Na+/K+-АТФаза (натрий-калиевая АТФаза или натрий-калиевый насос) содержится практически во всех плазматических мембранах и переносит ионы К+ в клетку, а ионы Na+ из клетки.
Nа+, К+ - насос - э то интегральный гликопротеид. Состоит из 4 субъединиц: двух α и двух β. β-субъединицы находится на наружной стороне мембраны и несут олигосахаридные цепи, α-субъединицы погружены в мембрану и несут участок, осуществляющий гидролиз АТФ, а также 3 участка связывания ионов Nа+ и два участка связывания ионов К+. α-субъединицы контактируют друг с другом и с β-субъединицами, а β-субъединицами пространственно разделены и друг с друго не контактируют (рис.64). Таким образом между субъединицами имеется некая полость. Этот насос переносит ионы Nа+ из клетки, а ионы К+ - в клетку. Так как, ионов Nа+ больше вне клеток, а К+ - внутри клеток, то транспорт с помощью насоса идет против градиента концентрации.
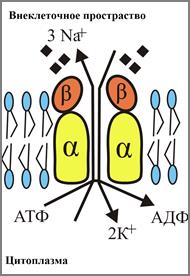
Рис. 64. Схема строения Nа+, К+-насоса
(Из: Мушкамбаров,2003)
Механизм действия насоса осуществляется следующим образом (рис. 65). В начале канал открыт внутрь клетки. 3 иона Nа+ связывают- ся с активными центрами связывания ионов Nа+ в АТФазе (1) и активируют фермент; он связывает молекулу АТФ катализирует её гидролиз, причем фосфатный остаток присоединяется к АТФазе (2). Происходит фосфорилирование фермента и он меняет свою конформацию: ионный канал закрывается с внутренней стороны мембраны и открывается с наружи (3). Ионы Nа+ в силу электрического отталкивания с помощью энергии гидролиза АТФ отсоединяются от центров связывания АТФазы и высвобождаются во внешнюю среду, несмотря на высокую их концентрацию здесь. Энергия гидролиза АТФ расходуется на фосфорилирование АТФазы и отсоединение ионов Nа+ от связывающих их центров в АТФазе.
Na+/K+-АТФаза, открытая с наружной стороны мембраныимеет специфические центры связывания 2 ионов К+ (4). Ионы К+ дефосфори- лируют АТФазу. При этом освобождается энергия АТФ, сохранившаяся в связи фосфатной группы.
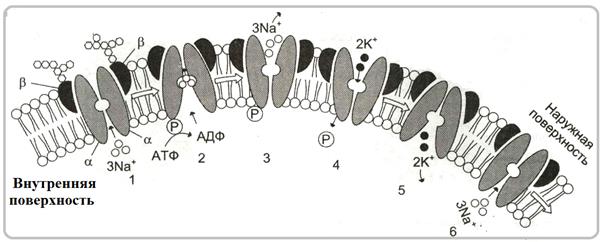
Рис. 65. Схема функционирования Nа+,К+-насоса.Объяснение в тексте
(Из: Северина,2007)
Дефосфорилирование изменяет конформацию АТФазы таким образом, что она возвращается в исходное состояние (5): ее полость вновь открыта внутрь клетки. Ионы К+ отсоединяются от фермента и освобождаются в цитозоль клетки. Так завершается цикл работы насоса. При этом из клетки удаляется 3 иона Nа+, а транспортируется внутрь два иона К+. Поскольку перенос катионов не равнозначен, то возникает разность электрических потенциалов. Из разности концентраций и разности электрических потенциалов клетки складывается трансмемб- ранный электрохимических потенциал.
Nа+, К+-насос необходим животным клеткам для поддержания осмотического баланса (осморегуляции). В клетке концентрация органи- ческих веществ выше, чем в межклеточном пространстве. Вследствие осмоса вода должна стремиться внутрь клетки. Клетка набухает, внутриклеточное давление увеличивается и происходит разрыв мембран (осмотический шок). Разность потенциалов создает препятствие для набухания клеток, уравновешивая избыток концентрации веществ.
Кроме того, он необходим клетке для поддержания электрической активности в нервных и мышечных клетках и, наконец, для активного транспорта некоторых веществ, например сахаров и аминокислот. Высокие концентрации калия требуются также для белкового синтеза, гликолиза, фотосинтеза и для некоторых других жизненно важных процессов.
- Na+-глюкозные ко-транспортёры (симпортёры) или Na+-зависимиый глюкозный насос (рис.66). Эти транспортёры обеспечивают активный транспорт глюкозы с помощью ионов Na+ и их работа зависит от градиента концентрации натрия, который формируется за счет работы Nа+, К+-насоса эпителиоцитов. Такие насосы работают только в почечных канальцах и кишечнике, обеспечивая всасывание глюкозы против градиента её концентрации.
Глюкоза через такие насосы проходит путем симпорта с ионами Na+ из фильтрата полости почечных канальцевв клетки их эпителия эпителиоциты. За счет энергии 1 молекулы АТФ в клетки транспор- тируется 3 иона Na+ и 3 молекулы глюкозы. Транспорт глюкозы из фильтрата в эпителиоциты обеспечивается энергией той АТФ, которая расходуется Nа+, К+-насосом эпителиоцитов. Из эпителиоцитов в кровь глюкоза транспортируется по механизму облегченной диффузии при участии траслоказы ГЛЮТ-2 из семейства ГЛЮТ (см. выше).
К активно транспортируемым через некоторые клеточные мембраны веществам относят ионы натрия, калия, кальция, железа, водорода, хлора, йода, мочевой кислоты, некоторые сахара и большинство аминокислот.
Нарушение работы ионных насосов сопровождается развитием патологических состояний в организме (так, необратимое повреждение клеток при недостатке кислорода связано с выключением транспортных АТФаз из-за отсутствия АТФ в условиях тканевой гипоксии).
Активный транспорт
Для эффективной работы некоторых клеток в определенных случаях необходимо, чтобы концентрация веществ внутри клеткибыла выше, чем во внеклеточной жидкости (например, ионы калия).Для других клеток, наоборот, необходимо сохранять концентрацию веществ внутри клетки на низком уровне, по сравнению с концентрацией вне клетки (например, ионы натрия). Например, вопреки уровню ее концентрации поглощается глюкоза; клетки щитовидной железы захватывают йод, при том, что его содержание в них в сотни раз выше, чем в крови; мышечная ткань выталкивает ионы натрия (при этом концентрация последних снаружи всегда выше, чем внутри) и накапливает ионы калия. Такие состояния не могут обеспечиваться простой диффузией, так как она в конечном итоге уравновесит концентрации этих ионов по обе стороны мембраны. Для создания избыточного движения ионов калия внутрь клетки, а ионов натрия — наружу необходим некий источник энергии. В этом случае транспорт веществ обеспечивается механизмом активного транспорта.
Активный транспорт – это перенос веществ через мембрану, который совершается против градиента концентрации (т.е. с компартмента с большой концентрации в компартмент с меньшей концентрацией) при участии специальных белков - транспортеров и требует затрат энергии АТФ, образующейся в процессе дыхания (рис.42). Поэтому в отсутствие дыхания активный транспорт идти не может. Энергия требуется для:
1 - преодоления естественного стремления вещества диф- фундировать по градиенту концентрации, то есть, в противоположном направлении;
2 - белки-транспортеры отличаются от траслоказ облегченной диффузии тем, что для обеспечения транспорта они должны изменять свою конформацию, для изменения которой и требуется энергия. На нужды активного транспорта расходуется примерно половина общих энергетических ресурсов клетки.
Активный транспорт может осуществляться по механизму унипорта (транспорт глюкозы в клетках печени), симпорта - сопряженного переноса (транспорт аминокислот или глюкозы вместе с ионами натрия в кишечных эпителиальных клетках, рис.66), либо антипорта (обмен ионов НСО3- на Cl- в мембране эритроцитов, рис.58).
Активным транспортом переносятся: 1 - минеральные ионы из межклеточной жидкости в клетку или обратно; 2 - аминокислоты из полости кишечника в клетки кишечника; 3 - глюкоза из первичной мочи в кровь др.