Гуморальная иммунная реакция осуществляется при участие В – лимфоцитов, макрофагов и Т – лимфоцитов (Т - хелперы), а также антигенов ГКГ – ІІ, расположенных на поверхности В –лимфоцитов, макрофагов, эпителиоцитов слизистой оболочки, эндотелиоцитов сосудов, тимуса и развивается параллельно с воспалительными процессами и клет.иммунным ответом.
Гуморальную реакцию вызывают растворимые антигены, корпускулярные антигены бактерий, органеллы, вводимые в клетку в изолированном виде и тд.
При слабовыраженной биол.агрессии для ее локализации и устранения бывает достаточно мобилизации механизмов первой линии защиты: воспаления и клет.иммунитета. Однако параллельно при нарастании биол.агрессии включается механизмы антигенспецифического гуморального иммунного ответа, которая развивается медленнее, но является более действенной и эффективной.
При первом пападении в организм антиген может быть опознан иммунной системой и способен индуцировать ответ лишь при условии:
1- его асязывания и обработкой антигенпредставляющими клетками (АПК). Это В – лимфоциты, активированные макрофаги и дендритные клетки лимфоидной ткани;
2- передаче информации о антигене от АПК лимфоидным клеткам в специализированных органах иммунной системы.
Антиген может быть доставлен в лимфоидные органы:
1 - растворимые с током жидкости (кровь или лимфа);
2- корпускулярные с помощью вспомогательных дендритных клеток.
При поступлении антигенного материала в лимфоидный орган происходит связывание и презентация антигена макрофагами и В – лимфоцитами. Раствоимая форма антигена взаимодействует с поверхностными Yg – рецепторами – В – клеток, для которых помощь АПК не нужна. При взаимодействии антигена с Yg – рецепторами В – клеток, последние узнают определенный специфический данным рецепторам участок антигена – эпитоп, после чего на мембране В – клетки образуется комплекс антиген - Yg – рецептор, который погружается внутрь клетки. Здесь эпитоп антигена взаимодействует с молекулами JKГ II мембраны В – клетки, образуя второй эпитоп, который будет распознаваться Т – хелперами, т.к. для последующего размножения, дифференцирован и и обеспечения иммунного ответа (синтеза антител) В – клеткам требуется активация Т – хелперами.
Корпускулярные антигены с вспомогательными клетками поглощаются макрофагами, подвергаются фагоцитозу и предварительной переработке: они фрагментируются до состояния антигенных детерминант, т.е. превращается в неполный антиген (гаптен). Затем гаптен выводится на поверхность В – клетки и соединяется с антигенами ГКГ – ІІ, образуя комплекс, в котором с помощью антигенов ГКГ – ІІ неполный антиген опять превращается в полный, но «стандартного» образца, становясь вторияно «корпускулярным», поскольку теперь он связан с поверхностью антиген представляющей клетки. Т.о., ферментируется «стандартный корпускулярный антиген» (СКА).
2. СКА опознается другими видами Т – лимфоцитов – Т хелпером и (помощником), которое по харектерному поверхностному белку называют еще СД4+ - клетками и различаются по иммуноспецифичности своих Т – клеточных рецепторов. Т – хелперы с помощью своих рецепторов узнают типовые черты чужеродных антигенов в составе СКА и взаимодействуют с ними. Это взаимодействие приводит к аетивации Т – хелперов: они подвергаются бласттрансформации в Т – зонах перифер.лимфоид. органов и дочерние клетки опять атакуют СКА, одновременно активируя В –клетки.
3. Активированное состояние В –клетки выражается в его бласттрансформации и активном размножением. Дочерные клетки вступают на путь дифференцировки в плазматические клетки плазмалоциты, которые накапливаются в лимфоузлах и селезенки, откуда выходят в кровь и мигрируют в ткани. Плазмациты продуцируют спецеф.антитела (иммуноглобулины), против чужеродных антигенных детерминант, вызывающих реакцию.
Антитела – это молекулы иммуноглобулинов, спецефически взаимодействующие с антигеноми и относящиеся к свевороточным иммуноглобулинам (в отличии от мембрано – связаным иммуноглобулинам (Yg) рецепторов В – клетки (BCR)). Они состоят из 3 ферментов: два Faв и один Fc – фрагмент. Faв – фрагмент связывается с антигеном, а Fc – фрагмент с макрофагом. Антитела составлят 5 классов иммуноглобулинов: YgМ, YgУ, YgА, YgД и YgЕ. Т.к. чужерод.антиген уничтожается растворимым антителом, то это реакция называется гуморальной.
Антиген образуется с антителом комплекс, который связывается своим Fc фрагментом с Fc – рецепторами на поверхности макрофагов (разновидность лейкоцитов) и нейтрофилов и фагоцитируется ими, либо антиген бактерий связывается с антителом и этот комплекс иницирует образование белков системы комплемента. Система комплемента – это около 20 белков плазмы крови, неактивные протеазы. Комплекс антиген – антитело активирует эти белки, которые каскадно активизируясь, присоединяется к антиген – антителу. Последний белок присоединяется к клеточной мембране, формируя в ней ионные каналы. В эти каналы устремляются протеазы, разрушающие внутриклет. белки и вода, способсвуя развитию осматического шока. Активация системы комплемента требует участия ионов Са2+ и Мg+.
При вторичном попадании антигена в клетку, он связывается с уже имеющимися против него антителами, которые сохраняются в крови и тканевой жидкости.
В процессе обеспечения иммунных реакций так же как при воспалении важную роль играют адгезивные взаимодействия. При гуморальном иммунитете межклеточные взаимодействие происходит: 1-при активации Т – хелпера антиген представляющей клеткой в составе СКА, т.е. клеткой макрофагом; 2 – при активации В – клетки Т – хелпером (табл.)
Оба типа межклеточных взаимодействий включают: 1 – иммуноспецифическое взаимодействие между рецепторами Т – хелпера и СКА; 2 – взаимодействие между адгезивными молекулами обеих клеток (имеется в виду Т – хелперы и В – клетки.)
1. в начале происходит иммунонеспецефическое взаимодействие макрофагов в комплексе с СКА с Т – хелперами с помощью Yg – подобного белка УСАМ – 1 макрофага и β2 – интегрина неактивного Т – хелпера (первичная адгезия). Если Т – клеточные рецепторы (ТСR) Yg-подоб.белок Т – клеток антиген ГКГ – ІІ комплементарны (иммуноспециф-е взаимодейстие), то запускается механизм усиления адгезии, после чего начинают секретироваться интерлейкины, ИЛ – I макрофагом и ИЛ – 2 Т – хелпером. ИЛ – 2 выделяется под влиянием ИЛ -1 и запускает механизм бластотрансформации Т – хелпера. Стимуляция клетки продуктом своей же секреции называется аутокринной. При активации Т – хелпера взаимодействия СКА – ТСR (2 по табл.) подкрепляется со стороны Т – хелпера еще двумя адгезивными белками СД3 и СД4, причем именно с СД4 первично связывается вирус СПИДа, проникаетв Т – хелпер, размножается с помощбю обратной транскриптазы (синтез ДНК по матрице РНК вируса), и разрушает Т – хелпер, что блокирует все гуморальные иммунные реакции,
Что и проявляется как Синдром Приобретенного ИммуноДефицита (т.е. СПИД).
2. Активная В – клеток активным Т – хелпером включает те же формы взаимодействия. Отличие заключается в том, что ИЛ -4 и ИЛ -5, которые индуцируют бласттрасформацию В – клеток (ИЛ -4) и стимулируют синтез и секрецию первичных антител класса YgM В клетками (ИЛ-5). Стимуляция В –клеток продуктами секреции клеток партнеров Т –хелперами называется гетерокринной.
При клеточном иммунитете взаимодействие между клетками – мишенями и Т –киллерами включает адгезивные взаимодействие между: 1) – УСАМ -1 (клетки -мишени) и λFA-1 (Т-киллеры); 2- СД58 (Кл.-мишени) и СД2 (Т-киллера); 3- узнавания антигена. ГКГ -1 и определение клетки как чужой в процессе взаимодействия между антигеном ГКГ -1 клетки-мишени и TCR клетки Т – киллера при поддерке белка Т – киллера СД8+ ; 4- взаимодействие между рецепторами белком FAs на поверхности клетки –мишени и FAs-лиганда (FAs-λ) на поверхности клетки Т –киллера. Это взаимодействие запускает апоптоз.
Межклеточные контакты
Межклеточные контакты – это различные виды соединений между клетками, образованные белками. Они обеспечивают непосредственную связь между клетками. Благодаря межклеточным контактам клетки взаимодействуют друг с другом на расстоянии с помощью сигналов, обусловленных сигнальными веществами (медиаторами), передаваемыми через межклеточное вещество.
Межклеточные контакты формируются в несколько фаз:
1 - узнавание одной клеткой другой клетки (может быть дистант- ными (через медиаторы)или контактным (при помощи рецепторов мембран);
2 - установление между клетками непрочных связей;
3- формирование устойчивых межклеточных контактов.
Вторая и третья фазы осуществляются при помощи молекул клеточной адгезии.
Различают следующие виды контактов (рис.89):
1- контакты простого типа;
2- контакты сцепляющего типа;
3- контакты запирающего типа;
4- контакты коммуникационного типа.
I. Контакты простого типа. Служат для соединения клеток. Различают простые межклеточные соединение и интердигитации или пальцевидные соединение.
Простые межклеточные соединение представляет собой адгезивное взаимодействие между мембранами соседних клеток. Здесь нет никаких дополнительных структур. Из адгезивных молекул взаимодействие обеспечивают кадгерины (у эпителиоцитов - это кадгерины Е и Р, у нервной и мышечной ткани – кадгерины N) и интегрины (рис.81,90).
Интердигитацин – при этом контакте цитоплазма одной клетки вклиниваются в виде пальца в цитоплазму другой клетки и наоборот. Поэтому его называют также пальцевидным соединением. Такое соединение увеличивает площадь межклеточных взаимодействий обеспечивая возрастание обмена веществ и усиливает прочность соединения. Встречается в тканях разного типа.
Контакты запирающего типа (прикрепительные контакты). Сюда относятся демосомы и адгезивный поясок, которые обеспечивают межклеточные взаимодействия и взаимодействия клеток с внеклеточным матриксом. В образовании таких контактов участвуют: филаменты цитоскелета, пластическая стр-ра, связывающиеся филаменты с мембраной и мембранные адгезивные белки.
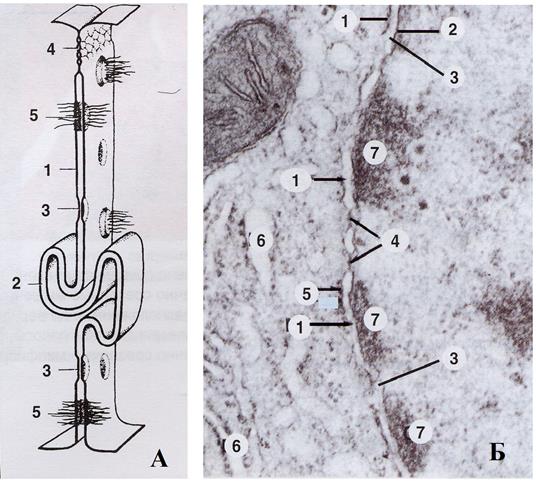
Рис. 89. Виды клеточных контактов –А – схематическое изображение, Б – электронная микрофотография
II.Контакты сцепляющего типа. К ним относятся десмосомы, полудесмосомы и адгезивный поясок.
Десмосомы (рис.) скрепляют клетки в отдельных местах в виде округлых образований, включающих мембраны обеих контактирующих плазмолемм. При этом с внутренней стороны этих мембран прилежит слой образованный белками десмоплакином и плакоглобином – цитоплазматическая пластинка. Который приклепляют промежуточные филаменты, отходящие в цитоплазму: в эпителиоцитах – кератины, в мышечной ткани – десмин, в мезихиме – виментин. Между контактирующими мембранами находятся утолщенный гликокаликс, содержащий сцепляющие белки – кадгерины: десмоглин и десмоколлин, связанные с белками цитоплазматической пластинки и выполняющие адгезивную функцию. На электронной микрофотографии выглядят как дисковидная пластинка, параллельная плазмолеммам.
Полудесмосомы – это контакты клеточной мембраны эпителиоцитов с базальной мембраной. Морфологически десмосема представляет половину десмосемы, обращенную к мембране дисковидной пластинкой. Функционально они облегчают взаимодействие клетки с внеклеточным матриксом. Дисковидная пластинка образована белком интегрином, а не кадгерином.
Количество десмосом и полудесмосом в контакте может быть несколько сотен. в случае нарушения десмосомного контакта (разрушение десмоглейна антителами) развивается пузырьчатка – смертельное инетическое заболевание.
Адгезивный контакт, образующийся между клетками имеющими несколько контактирующих поверхностей клетки однослойной эпителия контактируют с четырьмя такими же клетками.
Адгезивный поясок окружает клетку и связывает соединение клетки по строению сходен с десмосомой, но отличается по набору белков: 1- цитоплазмотическая пластинка образована белками катенинами, винкулином и λ актином; 2- от цитоплазм.пластинки отходят актиновые, а не промежуточные филаменты; 3- сцепляющие гентегральные адгезивные белки не десмоглеин и плакоглобин, а другие линкерные белки.
III.Контакты запирающего типа – плотные контакты. (рис) образуются между эпителиальными клетками. Основная особенность эпителиальных клеток заключается в том, что они служат барьерами обеспечивающими избирательный перенос в-в и исключающий любой промежуток между клетками. Эпителиальные клетки выолняют разнообразные важные функции, в том числе и функцию покроров клеток, плотно соединенных друг с другом и с базальной мембраной. Эти телиальные покровы образуют непроницаемый барьер между внешней и внутренней средой организма. Часто эпителий ткани испытывают нагрузку и для сохранения их целостности они должны остоваться плотно соединенными. У эпителия тонкого кишечника различают апикальную (с микроворсиками) мембрану, обращенную в полость кишечника, латериальные (боковые) мембраны и базальную мембрану, противоположную апикальной и соприкасающуюся с внеклет. Жидкостью и внеклеточным матриксом. Такое морфологич. Разделение обусловлено функцией эпителия кишечника: веасывания из просвета кишечника питат. Веществ и перенос их через соед.ткань в кровь. Молекулы поступающие в клетку из просвета кишечника через апикальную мембрану выносятся из нее через базальную мембрану. Плотные контакты препятствуют прохождению растворимых молекул между соседними клетками, плотно их смыкая. Образуется они интегральными адгезивными белками, но внешные их части не выступают над поверхностью плазмолемм, поэтому контактирующие мембраныпрактически вплотную прилегают друг-другу, опоясывая клетку в виде поясков, но не сплошных как адгезивные пояски, а ячеистые (т.е. местами мембраны остаются несомкнутыми, образуя ячеистую сеть).
IV. Контакты коммуникационного типа (щелевые). Через эти контакты осуществляется передача малых молекул из одной клетки в другую и называются они нексусы или щелевые контакты (рис)