Введение
Гуморальная составляющая межклеточных взаимодействий в иммунной системе опосредуется продуктами взаимодействующих клеток — цитокинами. Это белковые или полипептидные продукты активированных клеток иммунной системы, которые являются медиаторами межклеточных коммуникаций при иммунном ответе, гемопоэзе и развитии воспаления, эффекторами некоторых реакций иммунитета и служат связующим звеном между иммунной и другими системами организма. Значение цитокинов существенно выходит за рамки иммунологии, поскольку они играют важную роль в кроветворении, развитии патологии и т. д. Цитокины традиционно подразделяют на несколько групп:интерлейкины(факторы взаимодействия между лейкоцитами),интерфероны(цитокины с противовирусной активностью),факторы некроза опухолей(цитокины с цитотоксической активностью),колониестимулирующие факторы(гемопоэтические цитокины). Границы между группами условны.
По структуре выделяют несколько разновидностей молекул цитокинов. Подавляющее большинство из них в качестве характерного структурного элемента содержит 4 α-спирали и лишь для немногих (ИЛ-1, ФНОβ, трансформирующий фактор роста) характерно преобладание β-слоистой структуры.
Можно выделить 3 относительно автономные группы клеток — продуцентов цитокинов. Они характеризуются своим собственным типом ответа на активирующие воздействия и природой активаторов, а также собственным, хотя и значительно перекрывающимся набором продуцируемых ими цитокинов (табл. 1) и теми процессами, реализацию которых они обеспечивают. Это стромальные соединительнотканные клетки, которые вырабатывают цитокины и ответственны преимущественно за гемопоэз, моноциты/макрофаги, которые являются продуцентами цитокинов — медиаторов воспаления, и лимфоциты, вырабатывающие лимфокины, которые обеспечивают развитие антигенспецифической составляющей иммунного ответа.
В норме уровень продукции цитокинов стромальными клетками невысок. Стимулами для выработки этих цитокинов в отсутствие повреждающих и патогенных факторов служат, по-видимому, контакты с кроветворными клетками. Бактериальные продукты существенно усиливают выработку указанных цитокинов, причем это происходит не только в кроветворных органах, но и в очагах агрессии, что приводит к формированию экстрамедуллярных очагов кроветворения. В условиях активации аналогичную активность проявляют эпителиальные клетки кожи и слизистых оболочек.
Выработка цитокинов (монокинов) клетками миелоидно-моноцитарного происхождения индуцируется главным образом под влиянием бактериальных продуктов. Вызвать ее могут также многие метаболиты, сами цитокины, пептидные факторы, полиэлектролиты, а также контакты с окружающими клетками, процессы адгезии и фагоцитоза. Активация цитокиновых генов происходит в моноцитах и макрофагах в пределах 1 ч, и в ближайшие часы цитокин уже можно обнаружить в среде. Среди выделяемых этими клетками цитокинов преобладают факторы, участвующие в развитии воспаления. Их называют монокинами.
Третью группу клеток — продуцентов цитокинов (лимфокинов) составляют лимфоциты. Практически все разновидности лимфоцитов способны выделять цитокины, однако «профессиональными» продуцентами их являются СD4+-клетки-хелперы. Покоящиеся лимфоциты не продуцируют гуморальных факторов. Активация клеток осуществляется в результате связывания антигенраспознающих рецепторов и корецепторов. Самый ранний из лимфокинов — ИЛ-2 — появляется в цитоплазме Т-клеток через 2 ч после стимуляции; остальные лимфокины вырабатываются значительно позже и в определенной последовательности: ИЛ-4 через 4 ч, ИЛ-10 через 6 ч, ИЛ-9 через 24 ч. Пик выработки различных лимфокинов варьирует: 12 ч для ИЛ-2, 48 ч для ИЛ-4 и 5, 72 ч для ИЛ-9 и ИФНγ.
Таблица 1.Основные типы клеток — продуцентов цитокинов
| Клетки-продуценты | Индукторы цитокинов | Кинетика выработки | Продуцируемые цитокины |
| Стромальные клетки (фибробласты, эндотелиальные клетки) | Контактные взаимодействия, бактериальные продукты | В пределах часа мРНК, через 3-4 ч пик секреции цитокинов | ГМ-, Г-, М-КСФ; ИНФβ ИЛ-6,7,8,11 |
| Моноциты/макрофаги | Бактерии и их продукты, полиэлектролиты, форболовые эфиры | В пределах часа мРНК, через 6-14 ч пик секреции цитокинов | ИЛ-1,6; ФНОα ИЛ-10,12,15; ГМ-, Г-, М-КСФ, ТФРβ, ИНФα, хемокины. |
| Th1 | Связывание антигена/митогена через TCR-CD3/CD28+ИЛ-12 | Через 5-8 часов мРНК, через 10-48 ч пик секреции цитокина | ИЛ-2, ИНФγ, ФНОα и β, ИЛ-3, ГМ-КСФ, хемокины |
| Th2 | Связывание антигена/митогена+ИЛ-4 | Через 5-8 часов мРНК, через 24-48 ч пик секреции цитокинов | ИЛ-4,5,6,9,10,13,3; ГМ-КСФ, хемокины |
Примечание. ИНФ — интерферон, ТФР — трансформирующий фактор роста.
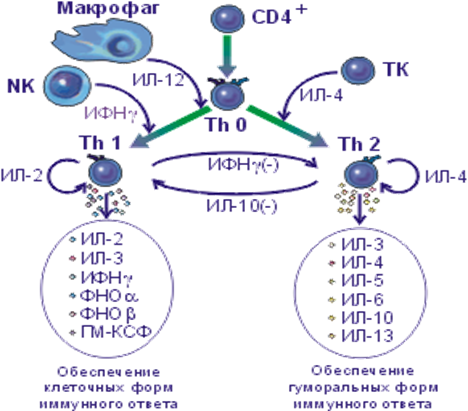
Эта последовательность отражает процессы дифференцировки Т-хелперов. «Наивные»СD4+-клетки в ответ на стимуляцию вырабатывают лишь ИЛ-2, затем, превращаясь в хелперы типа Th0, они начинают продуцировать в малом количестве широкий спектр цитокинов. При дальнейшей (или повторной) стимуляции Th0 дифференцируются на субпопуляции Тh1 и Th2, которые характеризуются различным спектром цитокинов (табл. 2), хотя и не различаются по мембранному фенотипу. Продукция цитокинов этими клетками значительно более высокая, чем в случае лимфоцитов, рассмотренных выше. Th1 вырабатывают ИЛ-2, ИФНγ, ФНОα и β, Th2 -ИЛ-4, 5, 6, 9, 10 и 13; оба типа клеток вырабатывают ИЛ-3 и ГМ-КСФ. В определенной степени аналогичная поляризация характерна также для СD8+-клеток, но интенсивность секреции цитокинов этими клетками значительно ниже, чем Т-хелперами.
Таблица 2.Типы Т-хелперов и их связь с системой цитокинов
| Тип Т-хелперов | Секретируемые цитокины | Ответ на действие цитокинов |
| Th0 | ИЛ-2, слабо — ИЛ-3,4,5,6,10,13, ИФНγ,ФНОα и β, ГМ-КСФ | ИЛ-12 и ИФНγ индуцирует развитие в направлении Тh1, ИЛ-4 и ТФРβ — в направлении Th2 |
| Th1 | ИФНγ, ИЛ-2, ФНОα и β, ИЛ-3, ГМ-КСФ, хемокины | ИЛ-2 - фактор роста, ИЛ-10 - ингибитор |
| Th2 | ИЛ-4,5,6,9,10,13,3; ГМ-КСФ, хемокины | ИЛ-2 и ИЛ-4 — факторы роста, ИФНγ и ИЛ-10 — ингибиторы |
Примечание. ИНФ — интерферон, ТФР — тромбоцитарный фактор роста.
Цитокины взаимосвязаны и образуют цельную систему взаимодействующих элементов - цитокиновую сеть. Для ее функционирования свойственны некоторые общие черты. Назовем некоторые из них.
Без антигенной стимуляции иммунной системы цитокиновая сеть функционирует на минимальном уровне. В отсутствие стимуляции клетки иммунной системы практически не выделяют цитокины и обычно не реагируют на них при их экзогенном введении. Синтез цитокинов и экспрессия их рецепторов, достаточная для развития ответа на эти факторы осуществляются при действии на клетки иммунной системы антигенов или иных стимулирующих агентов. Исключение составляют гемопоэтические цитокины, которые функционируют практически постоянно в ограниченных компартментах, а также некоторые цитокины, продуцируемые в малых количествах спонтанно. Такая особенность биосинтеза цитокинов связана с характером функционирования их генов. Практически все эти гены являются индуцибельными, т.е. для их активации требуется действие индукторов, в качестве которых выступают транскрипционные факторы, взаимодействующие с усиливающими (энхансерными) последовательностями регуляторного участка гена. Структура промоторныхучастков, ответственных за индукцию генов различных цитокинов и их рецепторов, различна, хотя и содержит общие элементы. Индукторами выработки цитокина и экспрессии его рецептора служат одни и те же факторы. Это обусловливает преимущественно локальный характер действия цитокинов. Локальность действия обеспечивается также тем, что экспрессия генов рецепторов продолжается дольше, чем экспрессия генов самих цитокинов. На уровне индивидуальной клетки экспрессия гена ИЛ-2 и выработка ИЛ-2 продолжаются дольше, чем экспрессия генов самих цитокинов. На уровне индивидуальной клетки экспрессия гена ИЛ-2 и выработка ИЛ-2 продолжается 1 сут, а экспрессия высокоаффинного рецептора - до 3 сут. За этот срок локально секретируемый цитокин успевает полностью утилизироваться клетками-мишенями. Кроме того, экспрессия рецептора усиливается под влиянием самого ИЛ-2. В результате местно секретируемый цитокин полностью потребляется в том микрообъеме, в котором он проявляет свое действие. В норме в кровотоке присутствуют следовые количества цитокинов недостаточные для проявления системных эффектов. Помимо указанных выше обстоятельств, это обусловлено чрезвычайно быстрым выведением цитокинов из кровотока через почки (время полужизни Т½составляет обычно минуты). Лишь у отдельных цитокинов время полужизни достаточно большое.
Для системы цитокинов характерна избыточность: каждая их разновидность может продуцироваться разными клетками (в связи с этим разделение их на моно- и лимфокины условно). Однако клетки одного и того же типа могут секретировать разные цитокины. Все цитокины полифункциональны, для них характерно значительное перекрывание функций. Цитокины могут усиливать или угнетать как выработку, так и функции друг друга. На рис. 1 представлен пример взаимосвязи между субпопуляциями Th1 и Th2, между которыми существует антагонизм. В регуляцию дифференцировки и соотношений этих клеток вовлекаются продукты нелимфоидных клеток, прежде всего макрофагов. В конечном счете это определяет структуру иммунного ответа и преобладающий характер иммунной защиты.
Таким образом, цитокины образуют систему (сеть) близкодействующих факторов, тесно взаимосвязанных на уровне продукции и осуществления эффекта. Цитокиновая сеть характеризуется высокой надежностью вследствие избыточности обеспечения основных влияний; условием ее функционирования служит активация клеток иммунной системы.
Интерлейкины
В настоящее время известны гены и установлены аминокислотные последовательности более двух десятков интерлейкинов (IL-1...IL-22), которые играют важную роль в формировании противоопухолевой защиты. При любом опухолевом росте имеются нарушения в системе интерлейкиновкоторые проявляются дисбалансом продукции и регуляции этих биологически активных веществ, изменением экспрессии соответствующих рецепторов.
Интерлейкины продуцируются различными клетками организма и являются факторами взаимодействия между клетками всех органов и систем. Во многих случаях они проявляют себя как факторы аутокринной регуляции.
Интерлейкин-1
Интерлейкин-1 (IL-1) - участвует практически во всех этапах иммуного ответа. Активирует APC и CD4 лимфоциты, влияет на дифференцировку Т- и В-лимфоцитов и других иммунокомпетентных клеток. IL-1 активирует цитотоксические Т-лимфоциты и NK-клетки, участвует в регуляции продукции IL-2, IL-4, IL-6, IL-8, гранулоцит-макрофаг колониестимулирующего фактора (GM-CSF) и других цитокинов. Активными ингибиторами продукции IL-1 являются IL-4, IL-10, IL-12, фактор некроза опухоли-альфа (TNF-α). Клонирование генов ИЛ-1 показало, что существует два различных белка с практически идентичной активностью — ИЛ-1α и β, которые имеют близкую молекулярную массу (15 000—17 000), но отличаются по изоэлектрической точке.
Основные продуценты ИЛ-1 — моноциты и макрофаги. ИЛ-1 образуется также В-лимфоцитами, белыми отростчатымиэпидермоцитами (клетками Лангерганса), глиальными, эндотелиальными и синовиальными клетками, фибробластами, эпителием кожи и тимуса, в культуре — некоторыми клонами Т-клеток. Условием выработки ИЛ-1 моноцитами и макрофагами является их активация бактериальными и иными продуктами (липополисахариды, некоторые экзотоксины, мурамилдипептид, пептидогликаны, митогены), а также вследствие адгезии, фагоцитоза. Синтез усиливается под действием цитохалахина В, колхицина, а также ингибитора синтеза белка циклогексимида (явление супериндукции). Ингибиторами синтеза ИЛ-1 являются простагландин Е2, глюкокортикоиды, факторы, повышающие уровень цАМФ.
Спектр клеток-мишеней ИЛ-1 чрезвычайно широк: активированные Т- и В-клетки, макрофаги, естественные киллеры, клетки эндотелия, мышц и хряща, базофилы, плазмоциты, кроветворные клетки. Все они экспрессируют рецепторы, общие для α- и β-форм ИЛ-1. Биологические эффекты ИЛ-1 можно условно разделить на иммунологические, воспалительные, кроветворные и межсистемные. ИЛ-1 причастен к запуску начальных событий иммунного ответа, в частности к вовлечению в него Т-хелперов. Среди Т-клеток рецепторы для ИЛ-1 в наибольшей степени экспрессируют «иммунные» Т-хелперы (Th2), продуцирующие ИЛ-3, 4 и 5, но не ИЛ-2 и интерферон у. Между тем ИЛ-1 способствует экспрессии генов ИЛ-2 и рецепторов для него. По-видимому, первый из этих эффектов реализуется посредством косвенных механизмов. Мембранный ИЛ-1 участвует в контактном взаимодействии Т-клеток с макрофагами. ИЛ-1 наряду с другими цитокинами вызывает пролиферацию активированных В-клеток и их дифференцировку в плазматические клетки. Введение ИЛ-1β вместе с антигеном способствует стимуляции антителообразования; более противоречивы данные о стимуляции гуморального иммунного ответа ИЛ-1α (возможно, он участвует в ограничении антителообразования).
ИЛ-1 стимулирует миелопоэз и ранние этапы эритропоэза. Действие ИЛ-1 в значительной степени связано с повышением выживаемости развивающихся клеток. С действием на кроветворение связан радиозащитный эффект ИЛ-1, проявляющийся при его введении до облучения и усиливающийся при введении через 5 сут после облучения. ИЛ-1 известен как провоспалительный агент. Он способен индуцировать большую часть местных и общих проявлений воспалительной реакции. Это достигается через повышение адгезивности эндотелия сосудов для клеток крови, увеличение прокоагулянтной активности клеток. ИЛ-1 повышает подвижность нейтрофилов, для ряда клеток является хемоаттрактантом, способствует активации клеток в очаге воспаления, усиливает продукцию ими других цитокинов, а также простагландинов, синтез коллагена и фибронектина, стимулирует фагоцитоз, генерацию супероксид-радикалов, вызывает дегрануляцию тучных клеток. Все это способствует развитию экссудативной и пролиферативной составляющих воспалительной реакции.
Таким образом, ИЛ-1, представленный двумя (а включая раИЛ-1 и тремя) молекулярными формами, является цитокином широкого спектра действия, продуцируемым преимущественно макрофагами. Он обусловливает пусковые реакции иммунитета, играет ключевую роль в развитии воспаления, участвует в регуляции гемопоэза, является медиатором взаимодействий между иммунной и нервной системами.
Интерлейкин-2
Интерлейкин-2 (IL-2) обладает выраженной способностью индуцировать активность практически всех клонов цитотоксических клеток. Он был первым интерлейкином, у которого была выявлена эта способность, и первым интерлейкином, который был применён Стивеном А. Розенбергом и сотрудниками для иммунотерапии рака.
IL-2 повышает цитолитическую функцию Т-киллеров и NK-клеток, увеличивает продукцию перфоринов и IFN-γ этими клетками, активирует моноциты и макрофаги, которые повышают синтез и секрецию TNF-α, IL-1β, IL-6, IL-8, гранулоцит-колониестимулирующего фактора (G-CSF), GM-CSF.
Секреция ИЛ-2 выявляется через 3—4 ч после стимуляции, достигает пика через 8—12 ч (раньше, чем секреция других лимфокинов) и прекращается через 24 ч. Invivo синтез ИЛ-2 достигает максимума через 1—3 сут после иммунизации и сохраняется в течение 12 сут.
На долю СD4+-лимфоцитов приходится 90% клеток-продуцентов ИЛ-2, на долю CD8+— около 10%. Основными продуцентами ИЛ-2 являются активированные Т-хелперы 1-го класса. Образование ИЛ-2 подавляют глюкокортикоиды (блокирующие активность гена ИЛ-2), оксимочевина, азатиоприн, ганглиозиды, дезоксиаденозин, а также простагландины и другие факторы, повышающие уровень цАМФ.
ИЛ-2 обладает относительно узким спектром мишеней и биологических эффектов. Основными его клетками-мишенями являются активированные Т- и В-лимфоциты и NK-клетки. Главное действие, оказываемое им на Т-лимфоциты, — индукция пролиферации в результате преодоления точки рестрикции между фазами цикла G1aи G1b. Присутствие ИЛ-2 необходимо и для осуществления дальнейших митозов. На этом основано получение длительных культур клонированных Т-лимфоцитов, хотя на определенном этапе они могут утрачивать чувствительность к ИЛ-2 (иногда при сохранении рецепторов). Возможно прямое активирующее действие ИЛ-2 без антигена или митогена, в этом случае требуются высокая концентрация ИЛ-2 и присутствие прилипающих клеток или ИЛ-1. ИЛ-2 предохраняет активированные клетки от апоптоза.
ИЛ-2 служит дифференцировочным фактором для Т-киллеров. Это действие ИЛ-2 проявляется позже, чем ростовое, и для его эффективного осуществления требуются дополнительные факторы, такие как ИЛ-6, 4, 7 и 12. ИЛ-2 способствует реализации функции Т-хелперов, усиливая выработку интерферона γ, экспрессию протоонкогенов и т.д. Он препятствует развитию иммунологической толерантности и даже отменяет ее.
Таким образом, ИЛ-2 представляет собой фактор роста и дифференцировки Т-лимфоцитов и NK-клеток, в меньшей степени В-лимфоцитов, продуцируемый активированными Т-хелперами. Он является важнейшим медиатором иммунитета (особенно клеточного) и участвует в реализации иммунной защиты и противоопухолевой резистентности.
Интерлейкин-3
Клетками — продуцентами ИЛ-3 являются Т-хелперы 1-го и 2-го классов, а также ряд других клеток (В-лимфоциты, миелоидные клетки, стромальные клетки костного мозга, астроциты головного мозга, кератиноциты). Активация гена ИЛ-3 наблюдается через 4 ч после стимуляции клетки и поддерживается несколько суток. Секреция ИЛ-3 выходит на плато через 16 ч и продолжается не менее 2 сут. Секреция ИЛ-3 подавляется циклоспорином А и глюкокортикоидами.
Клетками-мишенями ИЛ-3 служат в основном юные, в том числе полипотентные, кроветворные предшественники.
ИЛ-3 (в комбинации с сывороточнымикофакторами) обеспечивает поддержание пролиферации стволовых клеток и некомиттированных СD34+-предшественников;in vitroИЛ-3 обусловливает образование колоний смешанного и незрелого типов. В сочетании с линейно-специфическими факторами ИЛ-3 усиливает образование различных специализированных колоний кроветворных клеток, возможно, подготавливая кроветворные клетки к проявлению их действия.
Поскольку ИЛ-3 образуется в основном активированными Т-лимфоцитами, а не клетками стромы кроветворных органов, в которых синтез ИЛ-3 невелик, возникает вопрос о значении ИЛ-3 в регуляции гемопоэза. По-видимому, ИЛ-3 — преимущественно экстренный регулятор кроветворения, проявляющий свое действие при стрессе (когда Т-клетки мигрируют в костный мозг) и иммунном ответе, сопровождающемся активацией Т-клеток. In vivoИЛ-3 в большей степени усиливает экстрамедуллярный, чем костномозговой гемопоэз. ИЛ-3 влияет на ранние стадии В- и Т-лимфопоэза.
ИЛ-3 является ростовым фактором для тучных клеток слизистых оболочек (продуцирующих хондроитинфосфат) и усиливает ростстиму-лирующую активность ИЛ-4 в отношении тучных клеток соединительной ткани (серозные), усиливает продукцию ими гистамина. Таким образом, ИЛ-3 причастен к развитию аллергических реакций.
ИЛ-3 активирует эозинофилы и тормозит развитие NK-клеток. Следовательно, ИЛ-3 представляет собой продукт активированных Т-лимфоцитов, полипоэтин, влияющий в основном на ранние стадии гемопоэза и ответственный за его экстренную регуляцию. ИЛ-3 также участвует в развитии тучных клеток, подавляет формирование NK-клеток.
Интерлейкин-3 (IL-3) является полипотентным активатором гемопоэтических клеток. Роль IL-3 в опухолевом росте пока изучена недостаточно. Возможно его участие в противоопухолевой защите через стимуляцию NK-клеток и имеется синергизм с IL-4 в индукции активности некоторых CD4+лимфоцитов. IL-3 может усиливать опухолевуюцитотоксичность Т-лимфоцитов.
Интерлейкин-4
Интерлейкин-4 (IL-4) участвует в дифференцировке Т-хелперов: Th-0 в Th-1 и Th-2. Под действием IL-4 происходит переключение В-лимфоцитов на синтез IgE. IL-4 осуществляет контроль за регуляцией продукции TNF-α, IL-1β, IL-5, IL-6, IL-8, усиливает дифференцировку в цитотоксические Т-клетки, активирует макрофаги, усиливая их цитотоксический потенциал, индуцирует пролиферацию NK-клеток и при определенных условиях может участвовать в генерации LAK-клеток. Основными продуцентами IL-4 являются CD4+и CD8+лимфоциты, В-лимфоциты и макрофаги.
Главными продуцентами ИЛ-4 являются Т-хелперы 2-го класса (Тh2). При активации клеток экспрессия гена ИЛ-4 достигает пика через 6 ч, а синтез фактора — через 48 ч (т.е. позже, чем синтез ИЛ-2). Invitroмаксимум продукции ИЛ-4 наблюдается на 3-й сутки после иммунизации. ИЛ-4 синтезируется также тучными клетками и В-клеточными линиями. Продукция ИЛ-4 усиливается под влиянием ИЛ-1 и 2. Глюкокортикоиды в дозах, подавляющих синтез ИЛ-2, повышают образование ИЛ-4. Усиление синтеза ИЛ-4 происходит при старении.
Основными мишенями ИЛ-4 служат В-лимфоциты, для которых он является самым сильным ростовым фактором. Он действует на В-клетки раньше других ростовых факторов и может даже самостоятельно привести к активации и размножению покоящихся клеток. ИЛ-4 вызывает синтез аутокринных ростовых факторов В-лимфоцитами. ИЛ-4 усиливает выработку IgE и IgG1, вызывая переключение С-генов иммуноглобулинов. Он повышает экспрессию на В-лимфоцитах и тучных клетках и секрецию CD23 — низкоаффинного рецептора для IgE, который потенцирует выработку IgE. Эти эффекты, а также способность ИЛ-4 поддерживать пролиферацию серозных тучных клеток имеют прямое отношение к развитию аллергических реакций.
В отношении Т-лимфоцитов ИЛ-4 является как бы двойником-антагонистом ИЛ-2. ИЛ-4 вызывает пролиферацию тимоцитов (в сочетании с форболовым эфиром) и активированных зрелых Т-клеток, действуя сильнее на CD8+-, чем на СD4+-лимфоциты; среди последних на ИЛ-4 реагируют только Т-хелперы 2-го класса, т.е. его продуценты. ИЛ-4 является основным фактором, обусловливающим дифференцировку СD4+-клеток в направлении Th2. Он может выступать в роли дифференцировочного фактора и в отношении СD8+-клеток, способствуя образованию цитотоксических Т-лимфоцитов, которые вырабатывают набор цитокинов, свойственный Th2.
ИЛ-4 повышает экспрессию продуктов МНС II класса и антигенпрезентирующую активность вспомогательных клеток. В этом отношении ИЛ-4 является функциональным аналогом интерферона γ, хотя во многих других ситуациях он выступает как его антагонист. Прежде всего это проявляется в том, что ИЛ-4 и интерферон γ обусловливают альтернативные направления дифференцировки СD4+-клеток. Однако ИЛ-4 отменяет или ослабляет некоторые эффекты своих двойников — ИЛ-2 и интерферона γ, снижает синтез последних. Подавляя функции макрофагов и секрецию ими ИЛ-1, ФИО и ИЛ-6, ИЛ-4 оказывает противовоспалительное действие. В то же время он повышает цитотоксическую активность макрофагов, способствует миграции в очаг воспаления нейтрофилов, усиливает выработку колониестимулирующих факторов. ИЛ-4 стимулирует гемопоэз, вступая в кооперацию с другими факторами. В частности, он способствует выживаемости кроветворных клеток. Описано противоопухолевое действие ИЛ-4.
Таким образом, ИЛ-4 является главным продуктом Тh2-клеток, стимулирует их дифференцировку. Он обусловливает пролиферацию и дифференцировку В- и Т-лимфоцитов, влияет на развитие кроветворных клеток, на макрофаги, NK-клетки, базофилы, являясь функциональным двойником или антагонистом цитокинов, продуцируемых Тh1-клетками. ИЛ-4 способствует развитию аллергических реакций, обладает противовоспалительным и противоопухолевым действием.
Интерлейкин-5
ИЛ-5 описан как дифференцировочный фактор В-лимфоцитов. Почти одновременно было обнаружено его ростстимулирующее действие в отношении В-клеток, активированных декстран-сульфатом. Одно из его первоначальных наименований — «Т-замещающий фактор». ИЛ-5 продуцируется Т-хелперами 2-го класса. Экспрессия его гена и секреция проявляются относительно поздно после воздействия активаторов: максимальное накопление мРНК для ИЛ-5 в клетках-продуцентах происходит лишь к 3-м суткам.
Основные клетки-мишени ИЛ-5 — В-лимфоциты и эозинофилы. Эти клетки несут высоко- и низкоаффинные рецепторы для ИЛ-5, число которых на клеточной поверхности клетки значительно возрастает после активации. Способность ИЛ-5 поддерживать пролиферацию В-клеток реализуется на более поздних стадиях активации, чем действие ИЛ-4 и ИЛ-1. Это действие осуществляется в кооперации с ИЛ-2 (ИЛ-5 усиливает экспрессию рецепторов для ИЛ-2). ИЛ-5 способствует дифференцировке В-лимфоцитов в плазматические клетки, особенно в продуценты IgM. ИЛ-5 повышает образование IgA и менее значительно — синтез IgM, IgGl и IgE. Стимулируя выработку секреторногоIgA, ИЛ-5 способствует проявлению местной иммунной защиты слизистых оболочек.
ИЛ-5 является ростовым и дифференцировочным фактором эозинофилов; он способствует вовлечению этих клеток в воспалительные реакции, антипаразитарную и противоопухолевую защиту.
Таким образом, ИЛ-5 — продукт Тh2-клеток, обладающий относительно «узким» действием. Он является ростовым и дифференцировочным фактором В-лимфоцитов и эозинофилов, усиливает синтез IgA и тем самым способствует развитию местного иммунитета слизистых оболочек, стимулирует эозинофилы к участию в защите от паразитарных инвазий и опухолевого роста.
Интерлейкин-6
Клетки-продуценты ИЛ-6 чрезвычайно многочисленны: фибробласты, моноциты/макрофаги, лимфоциты, гепатоциты, кератиноциты, эндотелиальные, мезангиальные, кроветворные клетки, клетки трофобласта и опухолей различного происхождения. Условия индукции образования ИЛ-6 различны в зависимости от типа клеток — от простого прилипания к субстрату фибробластов и макрофагов до сложных событий активации лимфоцитов. Индукторами выработки ИЛ-6 могут быть бактериальные продукты, полиэлектролиты, митогены, а также ИЛ-1, ФНОα, интерфероны и колониестимулирующие факторы. Экспрессия мРНК для ИЛ-6 происходит в пределах 1 ч после активации, а секреция фактора — через несколько часов. Через 2 ч после внутривенного введения липополисахарида (ЛПС) уровень ИЛ-6 в сыворотке крови повышается в 1000 раз.
Не менее разнообразны клетки-мишени ИЛ-6. К ним относятся различные соединительнотканные элементы, клетки крови, иммунной и нейроэндокринной систем, печени. Высокоаффинный рецептор для ИЛ-6 образован двумя полипептидными цепями, одна из которых специфична для ИЛ-6, а другая (р130) является общей для группы цитокинов, выделяемой в семейство ИЛ-6. К этому семейству, помимо ИЛ-6, относятся ИЛ-11, онкостатин М, лейкозингибирующий фактор, кардиотрофин-1. Растворимая форма рецептора для ИЛ-6 обнаружена в моче.
Биологические эффекты ИЛ-6 сходны с таковыми ИЛ-1 и ФНОα. Прежде всего это участие в реализации воспалительной и иммунной реакций и кроветворения. При воспалении трудно разграничить эффекты названных цитокинов. По-видимому, ИЛ-6 более, чем два других флогогенных цитокина, влияет на синтез белков острой фазы гепатоцитами. Его действие на местные проявления воспаления аналогично действию ИЛ-1. ИЛ-6 способствует как обострению хронических, так и хронизации острых воспалительных процессов. Для него свойственно противовирусное действие. Выделяясь несколько позже, чем ИЛ-1 и ФНОα, ИЛ-6 подавляет их образование (они, наоборот, стимулируют его выработку) и поэтому относится к цитокинам, завершающим развитие воспалительной реакции.
В иммунной системе главной мишенью ИЛ-6 служат В-лимфоциты. ИЛ-6 является кофактором их пролиферации и самостоятельным дифференцировочным фактором. Он равномерно стимулирует выработку иммуноглобулинов всех классов. ИЛ-6 является, кроме того, фактором роста плазматических клеток и гибридом. При множественной миеломе он служит аутокринным фактором роста опухолевых клеток. В сочетании с другими факторами ИЛ-6 вызывает пролиферацию юных CD4-CD8--тимоцитов. Действуя на зрелые Т-клетки, ИЛ-6 подготавливает их к реакции на ИЛ-2, усиливает вызываемую ИЛ-2 пролиферацию Т-клеток и дифференцировку цитотоксических Т-лимфоцитов. Он повышает активирующее действие интерферона γ на NK-клетки и образование ЛАК-клеток под влиянием ИЛ-2.
На кроветворные клетки ИЛ-6 действует в основном как кофактор, способствуя проявлению эффектов ИЛ-3, ГМ-КСФ и М-КСФ. В результате ИЛ-6 усиливает образование invitroколоний всех типов. Не обладая самостоятельным радиозащитным действием, он повышает таковое ИЛ-1. ИЛ-6 выполняет определенную, пока недостаточно ясную роль во взаимодействии иммунной и нейроэндокринной систем.
Таким образом, ИЛ-6 представляет собой полифункциональный цитокин, продуцируемый фибробластами, макрофагами и другими клетками. По спектру биологического действия он близок ИЛ-1 и ФНОα, участвует в развитии воспаления, иммунных реакций, в регуляции кроветворения, служит ростовым фактором плазматических клеток, участвует в межсистемных взаимодействиях.
Интерлейкин-7
ИЛ-7 образуется стромальными клетками костного мозга и тимуса (фибробластами, эндотелиальными клетками, в тимусе также эпителиальными клетками), макрофагами.
ИЛ-7 представляет собой основной лимфопоэтин. Он участвует в коммитировании развития клеток крови в сторону В-лимфоцитов, является фактором выживания и роста проВ-клеток и ранних преВ-клеток, стимулирует пролиферацию внутритимусных предшественников Т-лимфоцитов (CD4-CD8--тимоцитов), включает реаранжировкуγ-генов TCR, усиливает экспрессию ингибитора апоптоза bcl-2, способствуя тем самым выживаемости преТ-клеток, обусловливает антигеннезависимое размножение Т-лимфоцитов вне тимуса. В качестве кофактора он влияет на пролиферацию Т-клеток и дифференцировку Т-киллеров; может индуцировать образование ЛАК-клеток. В составе рецептора для ИЛ-7 содержится γ-цепь (CD 132), общая для рецепторов ИЛ-2, ИЛ-4 и ряда других цитокинов.
В отличие от большинства других цитокинов, эффект которых дублируется, влияние ИЛ-7 на развитие лимфоцитов не имеет подобной «страховки», и удаление гена ИЛ-7 приводит
к опустошению тимуса, развитию тотальнойлимфопении и тяжелого иммунодефицита.
Итак, ИЛ-7, являясь лимфопоэтином, играет важную роль в обеспечении Т-клеточного звена иммунной защиты.
Интерлейкин-8
ИЛ-8, или фактор аттракции нейтрофилов (NAP-1), относится к группе хемоаттрактивных пептидов — α-хемокинов.
ИЛ-8 продуцируется многими типами клеток и обладает выраженными провоспалительными свойствами. Основным биологическим эффектом IL-8 является индукция хемотаксиса нейтрофилов, эозинофилов, базофилов и других клеток системы иммунитета. IL-8 усиливает ангиогенезinvivoиinvitro.
Интерлейкин-9
Интерлейкин-9 (IL-9) стимулирует выделение IL-2, IL-4, IL-6, IL-11, IFN-γ, IL-9 принимает участие в стимуляции цитотоксичности Т-киллеров и NK-клеток, индукции апоптоза. ИЛ-9 образуется в основном СD4+-Т-клетками типа Тh2 позже других цитокинов (экспрессия мРНК через 24 ч после стимуляции). Помимо основного эффекта — поддержания пролиферации активированных Т-хелперов — ИЛ-9 влияет на кроветворение (особенно на эритропоэз), активность тучных клеток и т.д.
Интерлейкин-10
Интерлекин-10 (IL-10) продуцируется Th-1 и Th-2, моноцитами, макрофагами и имеет широкий спектр действия с выраженным иммуносупрессорным эффектом. IL-10 снижает активность Th-1 в большей степени, чем Th-2. Противовоспалительная активность IL-10 проявляется способностью снижать продукцию провоспалительных цитокинов, усиливать продукцию антагониста рецептора IL-1 и уменьшать адгезию лейкоцитов к эндотелиальным клеткам, активированным IL-1. IL-10 может стимулировать синтез IgE. В своем ингибирующем действии на клеточный иммунитет IL-10 синергичен с IL-4.
Основной эффект ИЛ-10 заключается в подавлении синтеза цитокинов Thl-клетками (т.е. он обладает действием, противоположным влиянию интерферона у) и в снижении активности макрофагов, в том числе продукции воспалительных цитокинов. ИЛ-10 подавляет экспрессию молекул МНС II класса, пролиферацию Т-клеток, вызванную митогенами, а также развитие гиперчувствительности замедленного типа. В то же время ИЛ-10 выступает в качестве кофактора ИЛ-2 и ИЛ-7 в отношении пролиферации тимоцитов, служит синергистом ИЛ-4, усиливает пролиферацию В-клеток, защищает их от апоптоза, повышает синтез IgM и IgA. В результате ИЛ-10 способствует развитию гуморальной составляющей иммунного ответа, обусловливая антипаразитарную защиту и аллергическую реактивность организма.
Таким образом, ИЛ-10 служит важнейшим регулятором иммунного ответа, подавляющим активность макрофагов и Thl-клеток и обеспечивающим реализацию некоторых биологических эффектов Тh2.
Интерлейкин-11
Интерлейкин-11 (IL-11) провоспалительныйинтерлейкин, который регулирует функции Т- и В-лимфоцитов, принимает участие в индукции активности ряда киллерных клеток, является аутокринным фактором для пролиферации мегакариоцитов. Подобно IL-1 и IL-6 принимает участие в индукции синтеза белков острой фазы.
Он образуется фибробластами. ИЛ-11 обусловливает пролиферацию ранних кроветворных предшественников, подготавливает стволовые клетки к восприятию действия ИЛ-3, способствует миело- и эритропоэзу, развитию мегакариоцитов; однако может вызывать умеренную анемию. ИЛ-11 стимулирует иммунный ответ и развитие воспаления, способствуя дифференцировке нейтрофилов, выработке белков острой фазы, подавляет активность липопротеиновой липазы. По ряду эффектов ИЛ-11 может рассматриваться как функциональный двойник ИЛ-6 (в составе их рецепторов есть общая цепь — gpl30).
Интерлейкин-12
Интерлейкин-12 (IL-12) - полипотентный активатор клеточного иммунитета с противоопухолевой и антиметастатической активностью. Он усиливает активность Т-киллеров, NK- и LAK-клеток. IL-12 активирует и цитотоксичность макрофагов, а дефицит его продукции макрофагами может значительно снижать противоопухолевую активность. IL-12 оказывает противоопухолевый эффект, при раке легкого. Усиление роста опухоли, в частности, рака прямой кишки, ассоциируется со снижением продукции IL-12 и усилением продукции IL-10. Важным свойством IL-12 является усиление экспрессии FasL и индукция апоптоза. Рекомбинантный IL-12 способен препятствовать метастазированию в легкие и лимфатические узлы. Максимальный противоопухолевый эффект IL-12 наблюдается в сочетании с действием IL-2 и IFN-γ.
ИЛ-12 образуется макрофагами, дендритными клетками и В-лимфоцитами под влиянием стимуляции бактериальными продуктами. ИЛ-12 служит посредником между макрофагами и лимфоцитами, врожденным и приобретенным иммунитетом. Это проявляется в его способности регулировать соотношение клеточного и гуморального иммунного ответа через стимуляцию дифференцировки Т-хелперов в направлении Тh1. Этот эффект в значительной степени опосредован быстрым повышением продукции интерферона γ. ИЛ-12 стимулирует активность NK-клеток (естественных киллеров), пролиферацию этих клеток, обусловливает дифференцировку цитотоксических Т-лимфоцитов (оба эффекта осуществляются с участием интерферона γ); увеличивает образование ЛАК-клеток. С активацией Тh1- и NK-клеток связано противоопухолевое действие ИЛ-12 (его клинические испытания прошли успешно).
ИЛ-12 способс<