В главе о формировании половых клеток мы говорили, что ооциты к концу своего роста и созревания достигают состояния максимальной депрессии вследствие преобладания роста протоплазмы над ростом ядра, а также вследствие изменений последнего. Этим самым нарушаются нормальные, необходимые для правильного обмена веществ объемные соотношения между ядром и протоплазмой. В результате этого зрелое яйцо без оплодотворения обладает сравнительно короткой жизнеспособностью. Оплодотворение увеличивает вдвое объем яйцевого ядра и этим повышает обмен веществ в яйце, что проявляется в изменении характера окислительных процессов в нем. Но оплодотворением только частично восстанавливается нарушение соотношений ядра и протоплазмы, а именно компенсируется только то уменьшение ядра, которое произошло вследствие редукции хроматина при созревании.
Однако оплодотворение вызывает, кроме того, и дробление яйца, благодаря которому происходит дальнейшее восстановление нормальных соотношений между ядром и протоплазмой вследствие некоторых особенностей деления бластомер. Дело в том, что при обычном делении клетки и протоплазма и ядро последней делятся на две равные половины, так что получаются две клетки, в которых и ядро и протоплазма имеют половинные размеры по сравнению с материнской клеткой. Затем, благодаря ассимиляционной деятельности, плазма и ядро каждой из дочерних клеток растут и достигают размеров ядра и протоплазмы материнской клетки; вследствие этого соотношение ядра и протоплазмы в клетках во время деления остаются неизменными. При дроблении же, когда яйцо разделится на две бластомеры, ядро каждой из них вырастает приблизительно до размеров ядра яйца, по общий объем протоплазмы бластомер не изменяется, так как яйцо не может расти. Количество протоплазмы даже несколько уменьшается, так как часть ее объема занимается вырастающим ядром.
Так как такое же вырастание ядер при отсутствии роста протоплазмы происходит и при втором, и при всех последующих делениях дробящегося яйца, то с каждым делениембластомер соотношение ядра и протоплазмы изменяется в пользу ядра. Следовательно, изменение этих соотношений при дроблении идет в противоположном направлении по сравнению с теми изменениями, которые происходят во время роста ооцита, и соотношения ядра и протоплазмы с каждым делением приближаются все более к нормальным (Л ё б, Бовери, Годлевский). Теоретически (согласно гипотезе Лёба) можно предполагать, что при первом делении общее количество ядерного вещества в яйце должно возрасти вдвое, при втором делении вчетверо, при третьем в восемь раз и т. д.; однако, на самом деле наблюдаются некоторые отклонения от этих цифр. Так, в яйцах морского ежа (по Годлевскому) измерением диаметров отдельных ядер на последовательных стадиях и вычислением объемов ядер были получены следующие общие количества ядерного вещества на различных стадиях дробления (таблица 1).
Таблица 1
| Число делений | Стадии | Общие объемы ядер, µ3 | Геометрическая прогрессия (теоретически ожидаем.числа) |
| Женский пронуклеус Ядро дробления Стадия 2 бластомер 128 и след. | 1 300 1 582 2 084 19 938 20 262 30 747 | 1 300 2 600 5 200 44 600 83 200 166 400 |
В приведенном примере почти нет увеличения общего объема ядер после первого деления, и ожидаемый при нем общий объем ядер достигается только при втором делении. Увеличение ядер продолжается до шестого деления (стадия 64 бластомер), а начиная с седьмого деления, ядра с каждым делением уменьшаются и не растут, так что общий объем их не увеличивается до начала роста тканей зародыша.
Если сопоставить объемы ядра и протоплазмы в отдельных бластомерах и проследить изменения их соотношений при дроблении, то, по данным Рода-Эрдманн, у морского ежа получаются следующие цифры (таблица 2).
Таблица 2
| Стадии | Объем тканей клетки µ3 | Объем плазмы µ3 | Объем ядра µ3 | Отношение объема плазмы к объему ядра |
| 2 бластомер 64 – 132 Бластула | 100 000 43 000 22 343 9 351, 6 4 814 1 633 | 91 653 41 402 21 387 8 783 4 314 1 438 | 8 347 1 598 | 10,9 25,9 22,3 15,2 8,6 5,8 6,3 |
Эти цифры показывают, что уменьшение плазмо-ядерного соотношения, наблюдаемое во время дроблений, прекращается к шестому делению.
У пиявки Nephelis объем протоплазмы оогония увеличивается за время второго периода роста в 107 раз, а ядро ооцита, как мы указывали в свое время (стр. 16), не увеличивается совсем. В оогонии Nephelis соотношение объемов ядра и протоплазмы было 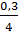 , а к концу второго периода роста
, а к концу второго периода роста  . Во время третьего периода роста оченьнемного уменьшается количество протоплазмы. При созревании яйца соотношение ядерно-плазменных объемов делается было
. Во время третьего периода роста оченьнемного уменьшается количество протоплазмы. При созревании яйца соотношение ядерно-плазменных объемов делается было 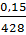 , но после оплодотворения оно снова становится равным . Так как при каждом делении дробления ядро в каждой бластомере дорастает до объема материнского ядра, т. е. объем ядер не меняется, а объем протоплазмы при этом уменьшается вдвое, то при первом делении объемное соотношение ядраи протоплазмы становится
, но после оплодотворения оно снова становится равным . Так как при каждом делении дробления ядро в каждой бластомере дорастает до объема материнского ядра, т. е. объем ядер не меняется, а объем протоплазмы при этом уменьшается вдвое, то при первом делении объемное соотношение ядраи протоплазмы становится  , при втором делении
, при втором делении 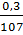 , при третьем делении
, при третьем делении 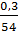 , причетвертом
, причетвертом  ,~, при пятом
,~, при пятом 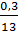 , при шестом
, при шестом  , при седьмом
, при седьмом 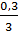 . Другими словами, соотношения объема ядра к объему протоплазмы к седьмому делению становятся приблизительно такими, какими они были в оогонии, до наступления его роста.
. Другими словами, соотношения объема ядра к объему протоплазмы к седьмому делению становятся приблизительно такими, какими они были в оогонии, до наступления его роста.
Конечно, изменения соотношении ядра и протоплазмы при дроблении не идут с математически ожидаемой точностью; после каждого деления ядра могут вырастать неодинаково, на некоторых делениях они совсем не вырастают, но зато могут усиленно вырастать па последующих делениях. Кроме того, соотношения объемов ядра и протоплазмы в отдельных бластомерах возвращаются не к соотношениям их в оогониях, а к соотношениям в клетках тела, которые, как правило, несколько отличаются в этом отношении от оогоний.
Кроме того, самый способ определения степени увеличения ядерного вещества, примененный Годлевским и состоящий в изменении общей массы этого вещества в определенном количестве яиц, которые находятся на одной и той же стадии, не может считаться достаточно точным. Несколько более точные данные получаются но методу Рода-Эрдманн измерением объема протоплазмы и ядра на следующих одна за другой стадиях развития.
Возврат к нормальным соотношениям ядра и протоплазмы в клетках в большинстве случаев наступает к шестому или седьмому делению яйца, но в яйцах некоторых животных это происходит несколько раньше, к пятому или даже четвертому делению, именно в тех случаях, когда в яйце существует видимая предварительная структура протоплазмы.
Так как яйцо является живой клеткой, то несомненно, что внутренняя физиологическая активность или внутренний метаболизм в нем или в его бластомерах на всех стадиях развития достаточно высок; но во время дробления этот метаболизм проявляется только внутри протоплазмы бластомер; по внешности же эти последние кажутся инертными, за исключением моментов деления клеток, сопровождающегося энергичными сокращениями протоплазмы. На стадиях дробления бластомеры еще не имеют той способности к взаимодействию, которая играет такую большую роль в позднейшем возникновении организации зародыша. Во многих случаях бластомера, изолированная от других бластомер, продолжает развиваться самодиференцировкой, т. е. проходит те же преобразования и дает те же производные, как и в нормальном окружении других бластомер. В своей группировке бластомеры могут очень слабо соприкасаться между собой и тогда легко отделяются друг от друга; если же соприкосновение между ними более тесно, то внешняя их инертность выражена в том, что в своей группировке они подчиняются действию поверхностного натяжения и располагаются так же, как группа капель жидкости в другой жидкости, не смешивающейся с ней и имеющей приблизительно одинаковый с ней удельный все. Эта внешняя пассивность бластомер является продолжением проявления той пассивности, которая свойственна яйцу до дробления, шаровидная форма которого также зависит от действия поверхностного натяжения.
Деление бластомер на стадиях дробления, как правило, происходит синхронно, и отклонения от этого правила объясняются или большим количеством желтка и неравномерным распределением его или рано выраженной во вне активностью некоторых бластомер. Синхронность дробления зависит не от связи бластомер друг с другом, а от одинаковых внутренних условий во всех бластомерах, так как деление остается синхронным и после искусственной изоляции их друг от друга.
Такое же отсутствие внешней активности отчасти сохраняется и в бластомерах стадии бластулы, вследствие чего форма последней также в значительной мере определяется поверхностным натяжением и в общем сходна у всех животных и ничего не говорит о морфологии будущего зародыша. Поэтому в морфологическом отношении не имеет оснований предположение Геккеля о повторении бластулой формы Volvox, так как и в том и в другом случае форма определяется внешней пассивностью или отсутствиемморфологических процессов. Этот первый период развития, т. е. дробление и, но крайней мере, часть стадии бластулы мы будем называть цитотипическим периодом, так как здесь видимые свойства стадий сводятся к внешним свойствам бластомер, вся активность которых сосредоточена на внутренних цитологических процессах.
Возврат ядерно-плазменных соотношений к норме обозначает возврат к нормальной интенсивности физиологической деятельности клеток и прекращение того состояния слабой внешней активности, в котором находилось яйцо со времени начала роста ооцита до конца дробления. Этот момент поэтому является очень важным, как изменяющий физиологию яйца; можно сказать, что им заканчивается дробление, и бластомеры становятся эмбриональными клетками. В морфологии яйца переход этот выражается началом морфологической диференцировки; в приведенном нами примере развития пиявки Nephelis после седьмого деления появляются гистологические отличия различных эмбриональных клеток и начинается гаструляция.
Второй период развития, в противоположность цитотипическому, можно назвать органотипическим. В этот период почти или вполне достигнуты нормальные и более или менее постоянные соотношения ядра и протоплазмы в эмбриональных клетках, так как сначала деление не сопровождается увеличением размеров ядра. Поэтому пропорционально уменьшается объем и ядра и протоплазмы, а позднее, когда начинается рост отдельных зачатков всего зародыша, ядро и протоплазма после каждого деления клетки растут пропорционально. Образование из крупных эмбриональных клеток большого количества мелких несколько улучшает условия обмена в каждой клетке, так как в мелких клетках отношения их поверхностей к объемам более благоприятны для обмена, чем в крупных клетках. Но в еще большей степени вследствие мельчания клеток улучшаются условия обмена в целых комплексах клеток, образующих зачаток того или другого органа, так как с уменьшением объемов клетки и увеличением их числа быстро увеличивается сумма всех поверхностей и протоплазмы и ядер этого комплекса в целом, а тем самым увеличивается и возможная поверхность обмена. Вместе с тем становится более тесным и контакт клеток зачатка как между собой, так и с клетками соседних зачатков. Тем самым отдельный зачаток приобретает характер отдельного многоклеточного участка организующегося целого, а весь зародыш—характер многоклеточного целого, так как создаются условия для физиологического взаимодействия и взаимной зависимости зачатков. Взаимодействие же клеток устанавливается потому, что с достижением нормальных ядерно-плазменных соотношений они перестают быть физиологически замкнутыми в себе бластомерами и получают возможность проявлять свои специфические свойства, различные в различных зачатках. Эти специфические свойства материала различных зачатков могут теперь проявляться и во вне, в морфологической диференцировке и в появлении определенной морфологии, характерной для зародыша данного типа животных. Поэтому только с началом органотипического периода изменения формы эмбрионального материала на различных стадиях получают значение для суждения о морфологии данного животного и о связи этой морфологии, с его филогенией.
ГЛАВА V. ДИФЕРЕНЦИРОВКА
Начальная диференцировка.Физиологические особенности начала оргапотипического периода вызывают начальную диференцировку органов, которая сопровождается появлением примитивной гистологической диференцировки. Последняя выражается в возникновении в клетках диференцирующегося органа или зачатка специфической структуры протоплазмы и в появлении в его клетках новых свойств, отличающих их от клеток других зачатков. В самом начале этих изменений, именно при диференцировке эктодермы и энтодермы при гаструляции, разница в характере клеток этих пластов возникает благодаря тому, что одни включения, имеющиеся в клетках бластулы, попадают исключительно в эктодерму, а другие — в энтодерму. Не менее важны наступающие при гаструляции изменения свойств эктодермы и энтодермы, которые обнаруживаются при некоторых экспериментах, показывающих различное отношение этих пластов к действию ионов. Если яйцо морского ежа со стадии дробления или бластулы поместить в морскую воду, в которой часть солей заменена изотопическим количеством солей лития, то при наступлении стадии гаструляции впячивание энтодермы не происходит, хотя в ней и проявлялись уже ее гистологические особенности, именно короткие мерцательные волоски, по сравнению с покрытой длинными мерцательными ресничками эктодермой. При этом оказывалось, что количество энтодермальных клеток заметно превышало их количество в нормальной гаструле, эктодермальных же клеток было значительно меньше нормального. Это отклонение от нормы было тем больше, чем больше находилось в растворе солей лития, так что в крайних случаях эктодерма была представлена маленькой кучкой клеток в виде пуговки на анимальном полюсе, а вся остальная стенка гаструлы составлялась из клеток энтодермы (рис. 19). Клетки мезенхимы оставались в нормальном своем количестве, так как находились в той же жидкости бластоцеля, как и в нормальной гаструле. Таким образом соли лития оказывались благоприятной средой для развития энтодермы и неблагоприятной для эктодермы. Морская вода с повышенным содержанием солей кальция и отсутствием аниона SO4, вызывала обратное действие, т. е. усиленное развитие эктодермы и подавление энтодермы. Такое же подавляющее действие соли лития оказывали и на эктодерму гаструлы лягушки, причем у этого объекта обратное влияние оказывал анион CNS(роданистых соединений). Следовательно, при переходе стадии бластулы в гаструлу в энтодерме и эктодерме возникают новые свойства, заставляющие их по разному реагировать на действие одного и того же внешнего реагента, наступает так называемая химическая диференцировка эмбрионального материала.
 Таким образом, в клетках энтодермы возникает потребность в иной благоприятной среде чем в клетках эктодермы, и, повидимому, сам процесс гаструляции, т. е. переход клеток будущей энтодермы в бластоцель, вызывается тем, что они нуждаются в той среде, которую они могут найти только в жидкости бластоцеля; для эктодермальных же клеток наиболее благоприятным условием является соприкосновениеих с внешней средой. Добавим еще, что но физико-химическим исследованиям гаструлы лягушки энтодерма обладает большой кислотностью, чем эктодерма.
Таким образом, в клетках энтодермы возникает потребность в иной благоприятной среде чем в клетках эктодермы, и, повидимому, сам процесс гаструляции, т. е. переход клеток будущей энтодермы в бластоцель, вызывается тем, что они нуждаются в той среде, которую они могут найти только в жидкости бластоцеля; для эктодермальных же клеток наиболее благоприятным условием является соприкосновениеих с внешней средой. Добавим еще, что но физико-химическим исследованиям гаструлы лягушки энтодерма обладает большой кислотностью, чем эктодерма.
Химическая диференцировка эктодермы и энтодермы по этим данным предшествует гаструляции и вызывает ее. Вместе с тем химическая диференцировка этих пластов дает и различное направление всей их дальнейшей диференцировке на различные производные, так как физико-химические и иные свойства, характеризующие энтодерму, ограничивают возможности изменений свойств клеток определенными рамками, в которых возможна только диференцировка в клетки либо кишечного типа, либо типа пищеварительных желез. Свойства же клеток эктодермы являются рамками, в которых возможно возникновение свойств только клеток наружных покровов и нервной ткани. Иногда эти возможности оказываются более обширными, и энтодерма дает кроме кишечника и его желез еще мезодерму и хорду (у позвоночных). Но это вопрос особый, связанный с вопросом о взаимной связи зародышевых зачатков, и мы остановимся на нем несколько дальше.
Дальнейшая диференцировка пластов на зачатки органов идет так, что каждый из них получает и химическую и начальную гистологическую диференцировку одновременно. Несомненно, что и морфологическое обособление органов имеет какие-то свои физиологические причины, но о них известно пока очень мало. Из определившихся уже в данной области вопросов необходимо отметить вопрос о взаимодействии различных зачатков, поставленный и в значительной мере разработанный Гольтфретером методом культивирования вне организма отдельных зачатков и различных комбинаций их. Из его исследований выяснено, что для правильной или организованной морфологической диференцировки зачатка требуется его соприкосновение с другим зачатком. Так, изолированный покровный слой в культуре не формируется в правильный эпителий, клетки ложатся беспорядочно, идет ненаправленный его рост. Но если с ним вместе культивируются клетки мезенхимы, последняя образует субстрат, на котором правильно формируется эпителий. Мезодерма правильно группируется и диференцируется лишь в том случае, если вместе с ней культивируется и хорда; в отсутствии контакта с хордой из эктодермы не может диференцироваться и нервная система и т. д. Организованная диференцировка возможна лишь при контактном взаимодействии различных зачатков, при котором, повидимому, изменяется проницаемость клеток и повышается интенсивность обмена веществ, так как находящийся в клетках желток начинает потребляться с повышенной скоростью. Очевидно, что такая связанность зачатков, которая ведет к образованию гармонически построенного организма, и характеризует зародыша на любой стадии как одно целое, как организм.
Другое явление, характеризующее начальную диференцировку, исследовано Чайльдом и названо физиологическим градиентом или градиентом восприимчивости и активности. При отравлении яйца или зародыша слабым раствором ядов (цианистый кали, формалин, сулема, щелочи и др.) умерщвление его идет постепенно и начинается в определенных точках его поверхности, откуда оно распространяется также в определенном направлении. Гибель этих участков можно легко констатировать, делая опыты отравления над прижизненно окрашенными яйцами, так как в отмерших участках краска исчезает. Причина большой чувствительности участка к яду лежит в его большей физиологической активности, следствием которой является более быстрое самоотравление клеток. Постоянно наблюдается повышенная активность переднего конца зародыша, от которого отравление распространяется постепенно назад, или анимального полюса яйца, откуда отравление распространяется к вегетативному полюсу. Ввиду того, что таким образом проявляется ступенчатая последовательность, направленная по главной оси зародыша или яйца, эта последовательность названа Чайльдом также осевым градиентом.
Такое же распределение восприимчивости и активности обнаруживается и в яйце до начала его дробления и является тогда выражением той предварительной структуры яйца, о которой неоднократно говорилось выше.
Кроме главного, или осевого, градиента, могут быть еще в некоторых яйцах и зародышах еще градиенты, идущие от определенной точки; например, в яйце лягушки одна точка повышенной активности находится на анимальном полюсе, а другая в определенном участке яйца ниже его экватора, откуда отравление распространяется к анимальному полюсу, навстречу осевому градиенту. Повышенная физиологическая активность много побочного участка служит признаком и большей активности в отношении образования новых производных, возникших в этом месте.
Существование одного главного градиента важно в том отношении, что от переднего (или анимального) участка зародыша зависит диференцировка органов позади лежащих частей (доминирование переднего конца), там как с него начинается диференцировка и создается та исходная организация, без которой не могут развиваться позади лежащие части тела. А этим в свою очередь характеризуется развитие всего зародыша, как единого целого.
Структурные изменения в клетках, или гистологическая диференцировка, не возникает с той же постепенностью, с какой идет размножение клеток. Зачаток органа возникает путем морфологической диференцировки, т. е. вследствие местного размножения клеток более общего зачатка делением, идущим в определенном направлении, но гистологически этот новый зачаток не отличается от исходного. Например, зачаток легких у позвоночных появляется в виде выпячивания стенки пищевода и сначала гистологически совершенно сходен с этим последним. Сохраняя то же гистологическое строение, он вырастает и начинает ветвиться. Но затем рост его временно приостанавливается, деление клеток прекращается, и все клетки зачатка одновременно и довольно быстро изменяют свою структуру, в то время как пищевод также изменяет структуру своих клеток, но иначе, чем зачаток легких. Деление клеток возобновляется с тем, чтобы снова остановиться, и тогда клетки опятьпретерпевают новую диференцировку, приближающую их структуру к структуре дефинитивного органа. Таким образом моменты новой гистологической диференцировки могут наступать не в связи с делениемклеток, а являются особым процессом, наступающим при морфологическом обособлении органа, но не совпадающим с ним во времени.
Морфологическая диференцировкаорганов возможна только при организованном развитии и не наступает, если в развитие искусственно внесены изменения, препятствующие гаструляции, которая является необходимым условием организованного развития. Но гистологическая диференцировкатканей может наступить даже при таких ненормальных условиях. Так, при культивировании яйца в полости тела животного или в его глазной орбите, гаструляции в нем не бывает, так как различия между внешней и внутренней средой, вызывающие процесс гаструляции, становятся в таких условиях слишком незначительными. Тем не менее через определенное время начинается гистологическая диференцировка, образуются типичные хрящевая, костная, нервная и мышечная ткани, но, несмотря на законченность своей диференцировки, все эти ткани не имеют определенной формы и располагаются совершенно хаотически в отношении друг к другу. Такая взаимная группировка их зависит повидимому от совершенно беспорядочных перемещений клеток в этих условиях.
С другой стороны, путем искусственных воздействий, например благодаря избытку КС1 в воде, Лилли удалось воспрепятствовать началу дробления яйца у Chaetopterus и Nereis, так что зародыш представлял собою не разделившееся на клетки целое даже к тому времени, когда нормально зародыш превращается в личинку. Но, несмотря на отсутствие деления плазмы и, следовательно, нормальной гистологической диференцировки, эта плазматическая масса морфологически была диференцирована приблизительно так, как нормальная личинка: там, где имеются в клетках нормальной личинки включения в виде определенного характера гранул, последние появляются и при отсутствии деления и т. д. Наконец, имеется довольно много яиц, в которых, если не гистологическая, то в некоторых отношениях ей отвечающая диференцировка протоплазмы, правда в несколько специфической форме, возникает до начала дробления в виде предварительной структуры яйца.
Образование и диференцировки органов в эмбриональной жизни не делает их способными функционировать у зародыша. Они не получают еще у зародыша законченную гистологическую диференцировку и только проделывают определенные фазы, которые к концу эмбрионального развития приводят к возникновению органов, способных к специальным функциям. Одни органы закладываются рано, но требуют продолжительного срока для своего развития, другие закладываются позже, так как развитие их идет сравнительно быстро, но только при выходе из яйца животного они способны функционировать. Та неполная диференцировка органов, которая обычно наблюдается у зародыша и не делает орган способным к выполнению его специальной функции, называется предфункциональной диференцировкой; последняя же степень диференцировки органа, наступающая обычно только к выходу из яйца и делающая орган способным к функции, называется функциональной диференцировкой. Правда, мускулатура тела на поздних стадиях развития зародыша уже может сокращаться, но эти сокращения мало полезны для зародыша, так как нервно-мышечный аппарат у них еще не готов и они показывают только, что способность к движению вследствие сложности необходимого для этого аппарата требует для возникновения настоящего двигательного акта некоторых предварительных фаз. Может быть, некоторую функциональную активность проявляют на поздних эмбриональных стадиях также пищеварительные железы кишечника, и в особенности печени.
Несомненно, однако, что все органы и части зародыша живут, так как только в таком случае они могут развиваться; следовательно, им во всяком случае свойствен обмен веществ, но этот обмен происходит не при помощи тех органов обмена, которые находятся в нем на стадиях морфологической диференцировки, так как на этих стадиях органы дыхания, пищеварения и выделения еще пользуются тем общим обменом веществ, который возможен в зародыше. Обычно при обмене поглощение кислорода и отдача углекислоты производятся всей поверхностью зародыша, питательные вещества — желток — перевариваются в тех клетках, которые его содержат, т. е. в большинстве случаев в клетках энтодермы, а затем в растворенном или ассимилированном виде диффузно распространяется и другие клетки. Выделение происходит во всех клетках, как это бывает и во взрослом организме, а продукты выделения уходят на ранних стадиях в бластоцель или в гастроцель, хотя, повидимому, в зародыше кроме углекислоты вообще мало образуется продуктов распада, так как желток представляет собой результат незаконченной ассимиляции белков данного животного, которая доводится до конца при развитии без образования значительного количества продуктов диссимиляции.
Преждевременная, или анахроничная, диференцировка. Из функций обмена функция выделения и функция транспортирования веществ по клеткам зародыша оказываются недостаточными для обслуживания всего зародыша. Поэтому у зародышей очень многих животных в помощь примитивному способу транспортирования и удаления веществ рано начинают функционировать специальные органы, как-то: эмбриональное сердце и эмбриональные органы выделения, причем последние служат не столько для удаления продуктов распада, которых мало, сколько для удаления из зародыша избыточной воды, т. е. для осморегуляции. Функциональная диференцировка таких органов наступает еще тогда, когда большая часть органов находится в состоянии предфункциональной диференцировки или еще раньше. Другими словами, эти органы достигают конечной диференцировки на сравнительно ранних стадиях развития всей системы органов, функционирующих у взрослого, и потому мы это явление называемпреждевременным, или анахроничным, функционированием или преждевременной диференцировкой. Она может возникать еще на ранних стадиях дробления, и, например, у дождевого червя Bimastus на стадии 4 бластомер две из них начинают функционировать как орган осморегуляции, и соответственно этому изменяется и специализируется их структура (рис. 88). Весь остальной зародыш возникает из других 2 бластомер этой стадии, что оказывается возможным благодаря некоторым специальным их особенностям, с которыми мы ближе познакомимся при описании развития аниелид.
В яйце паразитических гидромедуз Cuninа и Cunoctantha на стадии 2 бластомер одна преобразуется в амебообразную подвижную клетку (рис. 45), которая переносит другую бластомеру, дающую всего зародыша, из одной части тела матери в другую.
У иглокожих скелет вырабатывается клетками мезенхимы, поэтому у морских ежей н офиур, у которых сильно развитые скелетные иглы появляются еще у личинки, мезенхима, вырабатывающая их, появляется на стадии бластулы, тогда как у остальных она возникает только в конце стадии гаструлы со дна гастрального винпячения.
У некоторых брюхоногихмоллюсков на средней стадии развития, когда мезодерма, дающая сердце, далеко еще не сформирована, определенный участок эктодермы функционирует как сердце (рис. 113). У многих форм сначала наблюдается функционирование протонефридий, или личиночных органов выделения, которые затем сменяются метанефридиями, или дефинитивными органами выделения. В яйцах с большим количеством желтка широко распространена преждевременная диференцировка клеток, перерабатывающих желток и называемых желточной энтодермой, желточными клетками или вителлофагами.
У зародышей высших позвоночных на средних и ранних стадиях развития возникает в виде выроста задней части кишки пузыревидный орган — аллантоис, который покрывает почти вею поверхность желтка и функционирует как орган дыхания, так как обильно снабжен кровеносными сосудами, или (у млекопитающих) как орган передачи питательных веществ из матери в зародыш. Все эти органы относятся к так называемым провизорным органам, т. е. появляющимся у зародыша и исчезающим у взрослого животного; но их необходимо выделить как функционирующие провизорные органы, так как многие из последних, хотя и существуют, но не функционируют у зародыша (жаберные щели у высших позвоночных). Преждевременная диференцировка как видно из указанных примеров, наступает главным образом при возникновении различных эмбриональных приспособлений и является гистологической стороной их, которая одна и имеет значение в вопросах диференцировки.
Однако преждевременная диференцировкачасто имеет место не в каком-либо отдельном органе зародыша, а почти во всех эмбриональных зачатках; в таком случае она ведет к превращению зародыша в личинку, к которой уже трудно применять название эмбрионального приспособления. О значении преждевременной диференцировкипри метаморфозе будет сказано в соответствующей главе.
Приведенные примеры показывают, что потенциально самые разнообразные эмбриональные клетки, начиная с бластомер, могут быстро изменить свою структуру и достигнуть функциональной диференцировки. Но так как такая диференцировканаступает в клетках, имеющих еще совершенно эмбриональный характер, то подобный функционирующий орган имеет особое строение, не может войти в состав органов, функционирующиху взрослого животного, и прекращает свое существование к моменту выхода из яйца, а нередко и значительно раньше.
Некоторые особенно характерные для типа или класса органы, независимо от того, сохраняются они у взрослого животного или нет, рано приобретают функциональную диференцировку. Такова, например, хорда у хордовых: она рано диференцируется как у миноги, у которой она остается на всю жизнь, так и у высших позвоночных, у которых она почти совсем вытесняется дефинитивным скелетом. Для миноги она является органом с нормальной функциональной диференцировкой, для высших позвоночных она становится органом с преждевременной диференцировкой.
Первыми органами выделения у позвоночных диференцируются каналы предпочки в передних сегментах туловища, и для всех позвоночных последняя является органом с преждевременно диференцировкой. Затем после прекращения деятельности предпочки начинает функционировать первичная почка, занимающая среднюю и заднюю части туловища; у низших позвоночных она становится дефинитивным органом выделения, а у высших также в большей своей части исчезаеттак преждевременно диференцирующийся орган и заменяется вторичной почкой, функционирующей у взрослого.
Возможностью наступления такой преждевременной диференцировки особенно подчеркивается одна общая особенность хода эмбрионального развития, именно постепенность его и одинаковая степень эмбриональности всех клеток зародыша, из которых строятся органы готового животного. На стадиях дробления и бластулы бластомеры делятся синхронно и поэтому находятся на одинаковой ступени эмбриональиости, т. е. на каждой стадии одинаково удалены от состояния только что оплодотворенного яйца. В органотипический период деление имеет различные темпы в различных зачатках, но ускорение темпа ведет не к тому, что данный зачаток опережает другие в степени своего развития, а только к тому, что он дает или более крупный орган или является зачатком более сложным и разнообразным по своим производным. То же относится и к степени законченности развития и диференцировки на более поздних стадиях. В результате такого равенства в степени эмбриональности всех органов на различных стадиях развития зародыш появляется на свет с гармонично развитыми и одинаково достигшими состояния функциональной диференцировки органами.Преждевременная же диференцировка какого-нибудь эмбрионального зачатка выключает его из этой последовательной общей диференцировки органов зародыша.
Исключение из правила синхронности диференцировки представляет также развитие метамерных животных, т. е. обладающих осевой сегментацией тела с повторяющимися в каждом сегменте органами, но об этом придется сказать дальше по поводу всего вопроса о метамерных животных.
Кроме выключения клеток или зачатков из общего развития вследствие преждевременной диференцировки, клетки могут выключаться и вследствие задержки диференцировки некоторых из них. Таким образом возникают различные запасные клетки, имеющиеся в тех органах закончившего свое развитие животного, которые продолжают расти и функционально диференцироваться уже по окончании эмбрионального развития.
Запасные клетки также играют большую роль при восстановлении насильственно утерянных частей тела или регенерации и всегда имеются у животных, способных легко, регенерировать (см. главу X, «Регенерация»).
Гистологическая диференцировка, изменяя структуру протоплазмы клеток в сторону определенной специализации, лишает клетки возможности изменяться или затрудняет взаимное приспособление различных органов друг к другу. Такое приспособление, необходимое для гармоничного развития всей организации, возможно главным образом в моменты морфологической диференцировки органов, т. е. в начале их образования. Связывающее значение гистологической диференцировки важно в различных эмбриональных явлениях, и на него нам придется указывать и дальше. Данную же главу мы закончим рассмотрением одной особенности функциональной диференцировки, проявляющейся при развитии гибридов двух отдаленных друг от друга видов.
Если оплодотворить яйцо одного вида спермой другого, но принадлежащего к тому же роду вида, то развитие идет до конца и дает гибридов с признаками и отца и матери. Если же родители принадлежат к двум разным родам или семействам, то во многих случаях оплодотворение происходит и начинается развитие, которое, однако, дойдя при одних видовых комбинациях до одной стадии, при других комбинациях до другой, останавливается, появляются уродливые изменения зародыша, и последний вскоре погибает. Исследование причины гибели гибр