Триглециридов. По химическому строению триглицериды (или жиры) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот. С15H33,COOH-трипальмитин. Физико-хим. С-ва жиров во многом определяются составом жирных к-т. Жиры, содержащие преимущественно ненасыщенные жирные жирные к-ты, при комнотной комнотной температуре твердые, а ненасыщенные жирные к-ты- жидкие.Твердые жиры-животного происхождения,за исключением рыбьего жира. Жидкие-это раст.масла. Среди триглицеридов различают простые и смешанные. Первые являются сложными эфирами глицерина и одной из высших кислот.Вторые построены из остатка глицерина и остатков разных высших жирных кислот.
33.Ращепление жиров в ЖКТ. Расщепление жира на глицерин и высшие жирные кислоты осуществляется под влиянием фермента липазы. Для воздействия липазы на жир необходимо его предварительное эмульгирование, достигаемое путем перемешивания в кишечнике пищевой кашицы с желчью. В ротовой полости жиры не подвергаются химическим изменениям. В желудке присутствует липаза, однако ее активность невелика из-за отсутствия условий, необходимых для эмульгирования жира. В желудке гидролизуются только эмульгированные жиры — жиры молока и яичного желтка. В основном переваривание жира происходит в кишечнике и в первую очередь в двенадцатиперстной кишке, куда по протокам попадают вместе с желчью соли желчных кислот, обладающие мощным эмульгирующим действием. Желчные кислоты образуют тончайшую пленку на жировых каплях, которая препятствует слиянию отдельных капелек жира в более крупные капли. Это приводит к резкому увеличению поверхности соприкосновения жира с ферментом липазой и, следовательно, скорости гидролитического распада жира. К желчным кислотам относятся холевая, дезоксихолевая и другие. По своему строению они близки к холестерину. В желчи эти кислоты образуют с глицином (гликоколлом) или таурином парные соединения — глико- или таурохолевую, глико- или тауродезоксихолевую и другие желчные кислоты, присутствующие в виде натриевых солей. В клетках кишечного эпителия из продуктов гидролиза пищевых жиров вновь ресинтезируются жиры, или липоиды, специфичные для данного вида животных. Синтезированные липиды транспортируются в жировые депо. При необходимости из жировых депо жиры могут переходить в кровь и использоваться тканями в качестве энергетического материала.
35. Бета- окисленижир. Кис-т. Сущность бета-окисления заключается в том, что в ходе одного цикла хим. Превращений происходит окисление второго от-СООН группы атома углерода,который находитсяв бетаположении, и отщеплении молекулы ацетил-КоА. Далее ацетил-КоА вступает в цикл лимонной к-ты,затем в с-ме дыхательных ферментов окисляется до конечных продуктов-СО2 и Н2О с освобождением большого кол-ва энергии. Этот процесс протекает в митохондриях.Этот процесс проходит в 4 этапа: 1 окисление-дегидрирование заключается в отщеплении водорода от альфа и бета-атомов углерода в ацил-КоА при участии фермента ацил-КоА-дегидрогеназы, содержащий кофермент ФАД. 2-Гидратация-реакция присоединения молекулы воды по месту разрыва двойной связи под влиянием фермента еноилгидратазы с образованием гидроксиацил-КоА.Второе окисление сопровождается отщеплением двух атомов водорода от углеродного атома в бета-положении с участием НАД-содержащий дегидрогеназы.Тиолазная реакция-приводит к отщеплению ацетил-КоА от кетоацил-КоА при взаимодействии его с еще одной молекулой кофермента А. В р-те реакции образуется ацил-КоА и ацетил-Коа. Данный процесс катализируется ферментом тиолазой.
37 дыхательная цепь - система структурно и функционально связанных трансмембранных белков и переносчиков электронов. ЭТЦ позволяет запасти энергию, выделяющуюся в ходе окисления НАД∙Н и ФАДН2 молекулярным кислородом (в случае аэробного дыхания) или иными веществами (в случае анаэробного) в форме трансмембранного протонного потенциала за счёт последовательного переноса электрона по цепи, сопряжённого с перекачкой протонов через мембрану.
У прокариот ЭТЦ локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики расположены по своему окислительно-восстановительному потенциалу, транспорт электрона на всём протяжении цепи протекает самопроизвольно.
Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ. Сопряжённая работа ЭТЦ и АТФ-синтазы носит название окислительного фосфорилирования.
39. Патология углеводного обмена. Увеличение содержания глюкозы в крови — гипергликемия может происходить вследствие чрезмерно интенсивного глюконеогенеза либо в результате понижения способности утилизации глюкозы тканями, например при нарушении процессов ее транспорта через клеточные мембраны. Понижение содержания глюкозы в крови — гипогликемия — может являться симптомом различных болезней и патологических состояний, причем особенно уязвимым в этом отношении является мозг: следствием гипогликемии могут быть необратимые нарушения его функций.
Генетически обусловленные дефекты ферментов У. о. являются причиной многих наследственных болезней. Примером генетически обусловленного наследственного нарушения обмена моносахаридов может служить галактоземия, развивающаяся в результате дефекта синтеза фермента галактозо-1-фосфатуридилтрансферазы. Признаки галактоземии отмечают также при генетическом дефекте УДФ-глюкоза-4-эпимеразы. Характерными признаками галактоземии являются гипогликемия, галактозурия, появление и накопление в крови наряду с галактозой галактозо-1-фосфата, а также снижение массы тела, жировая дистрофия и цирроз печени, желтуха, катаракта, развивающаяся в раннем возрасте, задержка психомоторного развития. При тяжелой форме галактоземии дети часто погибают ни первом году жизни вследствие нарушений функций печени или пониженной сопротивляемости инфекциям.
2.моносахариды. строение, изомер…. Моносахариды, строение, изомерия, свойства
Моносахариды можно рассматривать как производные многоатомных спиртов, содержащие карбонильную группу

| и | 
|
Если карбонильная группа находится в конце цепи, то моносах альдегидоспирт и называется альдозой; при любом другом положении этой карбонильной группы моносахарид является кетоспиртом и называется кетозой.
По количеству атомов углерода различают триозы, тетрозы, пентозы, гексозы, гептозы.
Простейшим представителем моносахаридов являются триозы: глицериновый альдегид (альдоза) и дигидроксиацетон (кетоза).
При окислении первичной спиртовой группы трехатомного спирта – глицерина образуется глицериновый альдегид (глицеральдегид), а окисление вторичной спиртовой группы приводит к образованию дигидроксиацетона.

глицерин альдегидоглицерин дигидроксиацетон эритроза (тетроза) рибоза (пентоза) глюкоза (гексоза) фруктоза (гексоза)
Для моносахаридов характерна оптическая изомерия. Все моносахариды содержат один или более асимметрических атомов углерода: у альдозы - 1; у альдотетрозы - 2; у альдокетозы - 3; у альдогексозы - 4. 
Кетозы с тем же числом атомов углерода в молекуле содержат на один асимметричный атом меньше. Общее число стереоизомеров для любого моносахарида выражается формулой: N = 2n, где N - число стереоизомеров; n - число асимметричных атомов углерода. Глюкоза имеет 16 стереоизомеров.
Тот изомер, у которого при проекции молекулы на плоскость группа - ОН у асимметричного атома углерода расположена с правой стороны, принято считать D-изомером, а его зеркальное отражение - L-изомером. Например,

D-изомер L-изомер
Химические свойства моносахаридов (на примере глюкозы)
· Глюкоза способна подвергаться спиртовому брожению с образованием СО2 и спирта: С6Н12О6 2 С2Н5ОН + 2СО2
· Глюкоза в организме человека может окисляться:
а) В присутствии кислорода: С6Н12О6 +6О2 +36Н3РО4 +36АДФ → 6СО2 + 42 Н2О + 38АТФ;
б) В гипоксических условиях - с образованием молочной кислоты
С6Н12 О6 = 2СН3СНОНСООН + 2АТФ.
· Фосфорилирование глюкозы:

глюкоза фосфорная к-та глюкозо-1-фосфат
· Моносахариды легко окисляются, т. к. содержат карбонильную группу:
а) реакция серебряного зеркала
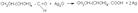
глюкоза глюконовая к-та
б) реакция Троммера:

глюкоза глюконовая кислотата
в) реакция с реактивом Фелинга:

глюкоза + реактив Фелинга = глюконовая к-та + натрий-калиевая соль винной кислоты
5. Восстановление моносахаридов:

глюкоза (альдегидо-спирт) сорбит (шестиатомный спирт)
4. общая хар-каБелки- важнейшая частьпищи и суточ потребность взрослого 1,5-2 г на 1кг. Велико значение в дет питании 5-6 на 1 кг. В сотав вход22ам-ты, (8незамен).Белковая молекула имеет четыре уровня организации: первичная, вторичная, третичная и четвертичная.
Первичная структура белковой молекулы представляет собой определенную последовательность расположения аминокислотных остатков, которые объединяются пептидными связями. Её устойчивость обеспечивается наличием водородных связей.
Вторичная структура белка определяется расположением полипептидной цепочки в пространстве в виде спирали или складчатой формы, которая удерживается водородными связями.
Третичная структура молекулы белка представляет собой более сложную конфигурацию и упаковку полипептидной цепочки в виде глобулярной (шарообразной) или фибриллярной (нитевидной) формы. В ее формировании принимают участие ионная, водородная связи и гидрофобные взаимодействия.
Четвертичная структура - это соединение глобул в комплексы между собой или связь субъединиц с веществами небелковой природы. В образовании четвертичной структуры белка принимают участие слабые нековалентные связи. Четвертичную структуру имеют многие ферменты, гемоглобин, миозин, миоглобин и др.
6. Общая хар-ка нуклеиновых…. Дезоксирибонуклеи́новая кислота́ (ДНК) — макромолекула (одна из трех основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. Основная роль ДНК вклетках — долговременное хранение информации о структуре РНК и белков. Рибонуклеи́новая кисло́та (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.А-Т,Г-Ц—ДНК,Рнк---А-У,Т-Ц.Роль Днк-хранения и передача наследственной информации.Роль Рнк-синтез белков по программе зашифрованной в участках ДНК.
8. Триглециридов. По химическому строению триглицериды (или жиры) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот. С15H33,COOH-трипальмитин. Физико-хим. С-ва жиров во многом определяются составом жирных к-т. Жиры, содержащие преимущественно ненасыщенные жирные жирные к-ты, при комнотной комнотной температуре твердые, а ненасыщенные жирные к-ты- жидкие.Твердые жиры-животного происхождения,за исключением рыбьего жира. Жидкие-это раст.масла. Среди триглицеридов различают простые и смешанные. Первые являются сложными эфирами глицерина и одной из высших кислот.Вторые построены из остатка глицерина и остатков разных высших жирных кислот
10. Предмет биохимии ….Биохи́мия — наука о химическом составе живых клеток и организмов и о химических процессах, лежащих в основе ихжизнедеятельности. Термин «биохимия» эпизодически употреблялся с середины XIX века, в классическом смысле он был предложен и введён в научную среду в 1903 году немецкимхимиком Карлом Нойбергом. Как самостоятельная наука биохимия сформировалась примерно 100 лет назад, однако биохимические процессы люди использовали ещё в глубокой древности, не подозревая, разумеется, об их истинной сущности. Биохимия стала первой биологической дисциплиной с развитым математическим аппаратом благодаря работам Холдейна, Михаэлиса, Ментен и других биохимиков, создавших ферментативную кинетику, основным законом которой является уравнение Михаэлиса-Ментен.Открытие ферментов позволило начать грандиозную работу по полному описанию всех процессов метаболизма, не завершённую до сих пор. Одними из первых значительных находок в этой области стали открытия витаминов, гликолиза и цикла трикарбоновых кислот.
12. Особенности обменя вещ-в. Цереброзиды содержат спирт сфингозин,жирные кислоты и остатки различных сахаров.Содержатся в клетках белого вещества мозга,присутствуют в мембранах других клеток.Ганглиозиды-это наиболее сложные по строению сфинголипиды.В их состав входит несколько остатков сахаров.Они содержаться в сером веществе мозга.. Макроэргические соединения — группа природных веществ, молекулы которых содержат богатые энергией, или макроэргические, связи; присутствуют во всех живых клетках и участвуют в накоплении и превращении энергии. Разрыв макроэргических связей в молекулах М.с. сопровождается выделением энергии, используемой для биосинтеза и транспорта веществ, мышечного сокращения, пищеварения и других процессов жизнедеятельности организма. Все известные М.с. содержат фосфорильную (—РО3Н2) или ацильную группы и могут быть описаны формулой Х—Y, где Х — атом азота, кислорода, серы или углерода, а Y — атом фосфора или углерода. Реакционная способность М.с. связана с повышенным сродством к электрону атома Y, что обусловливает высокую свободную энергию гидролиза М.с., составляющую 6—14 ккал/моль. Аденозинтрифосфа́т — нуклеотид, играет исключительно важную роль в обмене энергии и веществ в организмах; в первую очередь соединение известно как универсальный источник энергии для всех биохимических процессов, протекающих в живых системах. Главная роль АТФ в организме связана с обеспечением энергией многочисленных биохимических реакций. Являясь носителем двух высокоэнергетических связей, АТФ служит непосредственным источником энергии для множества энергозатратных биохимических и физиологических процессов. Все это реакции синтеза сложных веществ в организме: осуществление активного переноса молекул через биологические мембраны, в том числе и для создания трансмембранного электрического потенциала; осуществления мышечного сокращения.
14. Ферме́нты — обычно белковые молекулы или молекулы РНК (рибозимы) или их комплексы, ускоряющие (катализирующие) химические реакции в живых системах. Реагенты в реакции, катализируемой ферментами, называются субстратами, а получающиеся вещества — продуктами. Ферменты специфичны к субстратам. Ферментативная активность может регулироваться активаторами и ингибиторами (активаторы — повышают, ингибиторы — понижают). Белковые ферменты синтезируются на рибосомах, а РНК — в ядре. Структура и механизм действия ферментов Как и все белки, ферменты синтезируются в виде линейной цепочки аминокислот, которая сворачивается определённым образом. Каждая последовательность аминокислот сворачивается особым образом, и получающаяся молекула (белковая глобула) обладает уникальными свойствами. Несколько белковых цепей могут объединяться в белковый комплекс. Третичная структура белков разрушается при нагревании или воздействии некоторых химических веществ.Чтобы катализировать реакцию, фермент должен связаться с одним или несколькими субстратами. Белковая цепь фермента сворачивается таким образом, что на поверхности глобулы образуется щель, или впадина, где связываются субстраты. Эта область называется сайтом связывания субстрата. Обычно он совпадает с активным центром фермента или находится вблизи него. Некоторые ферменты содержат также сайты связывания кофакторов или ионов металлов.Фермент, соединяясь с субстратом:очищает субстрат от водяной «шубы» располагает реагирующие молекулы субстратов в пространстве нужным для протекания реакции образом
подготавливает к реакции (например, поляризует) молекулы субстратов. Обычно присоединение фермента к субстрату происходит за счет ионных или водородных связей, редко – за счет ковалентных. В конце реакции её продукт (или продукты) отделяются от фермента.В результате фермент снижает энергию активации реакции. Это происходит потому, что в присутствии фермента реакция идет по другому пути (фактически происходит другая реакция), например:В отсутствии фермента:А+В = АВ В присутствии фермента:
А+Ф = АФ АФ+В = АВФ АВФ = АВ+Ф где А, В - субстраты, АВ - продукт реакции, Ф - фермент. Свойства: они остаются неизмененными после реакции, т.е. освобождаясь, могут вновь реагировать с новыми молекулами субстрата. Ферменты способны оказывать действие в ничтожно малых концентрациях
16. .Общая хар-ка углеводов. Углево́ды(сахара, сахариды) —органические вещества, содержащиекарбонильную группуи несколькогидроксильных групп. Название класса соединений происходит от слов «гидраты углерода», оно было впервые предложено К. Шмидтом в1844 году. Появление такого названия связано с тем, что первые из известных науке углеводов описывались брутто-формулой Cx(H2O)y, формально являясь соединениямиуглеродаиводы.Моносахари́ды(от греческогоmonos — единственный,sacchar —сахар) — простейшие углеводы, не гидролизующиеся с образованием более простых углеводов — обычно представляют собой бесцветные, легко растворимые вводе, плохо — в спирте и совсем нерастворимые в эфире, твёрдые прозрачныеорганические соединения[2], одна из основных групп углеводов, самая простая форма сахара.Дисахари́ды— сложныеорганические соединения, одна из основных групп углеводов, пригидролизекаждая молекула распадается на две молекулы моносахаридов. О́лигосахари́ды — углеводы, молекулы которых синтезированы из 2 — 10 остатков моносахаридов, соединённых гликозидными связями. Структурная и опорная функции. Углеводы участвуют в построении различных опорных структур. Так целлюлоза является основным структурным компонентом клеточных стенокрастений, хитин выполняет аналогичную функцию у грибов, а также обеспечивает жёсткость экзоскелета членистоногих[1].Защитная роль у растений. У некоторых растений есть защитные образования (шипы, колючки и др.), состоящие из клеточных стенок мёртвых клеток.Пластическая функция. Углеводы входят в состав сложных молекул (например, пентозы (рибоза и дезоксирибоза) участвуют в построении АТФ, ДНК и РНК)..Энергетическая функция. Углеводы служат источником энергии: при окислении 1 грамма углеводов выделяются 4,1 ккал энергии и 0,4 г воды.Запасающая функция. Углеводы выступают в качестве запасных питательных веществ: гликоген у животных, крахмал и инулин — у растений.Осмотическая функция. Углеводы участвуют в регуляции осмотического давления в организме. Так, в крови содержится 100—110 мг/% глюкозы, от концентрации глюкозызависит осмотическое давление крови.Рецепторная функция. Олигосахариды входят в состав воспринимающей части многих клеточных рецепторов или молекул-лигандов.
18. Дисахариды и полисахариды. Строение и свойства, биолог.родь. Представители. Дисахариды - типичные сахароподобные полисахариды. Это твердые вещества, или некристаллизующиеся сиропы, хорошо растворимые в воде. Как аморфные, так и кристаллические дисахариды обычно плавятся в некотором интервале температур и, как правило, с разложением. Дисахариды образуются в результате реакции конденсации между двумя моносахаридами, обычно гексозами. Связь между двумя моносахаридами называют гликозидной связью. Обычно она образуется между первым и четвертым углеродными атомами соседних моносахаридных единиц (1,4-гликозидная связь). Этот процесс может повторяться бессчетное число раз, в результате чего и возникают гигантские молекулы полисахаридов. После того как моносахаридные единицы соединятся друг с другом, их называют остатками. Таким образом мальтоза состоит из двух остатков глюкозы. Среди дисахаридов наиболее широко распространены мальтоза (глюкоза + глюкоза), лактоза (глюкоза + галактоза), сахароза (глюкоза + фруктоза).
ОТДЕЛЬНЫЕ ПРЕДСТАВИТЕЛИ ДИСАХАРИДОВ. Мальтоза (солодовый сахар) имеет формулу С12Н22О11. Название возникло в связи со способом получения мальтозы: ее получают из крахмала при воздействии солода (лат. maltum - солод). В результате гидролиза мальтоза расщепляется на две молекулы глюкозы: С12Н22О11 + Н2О = 2С6Н12О6)Лактоза (молочный сахар). Название этого дисахарида возникло в связи с его получением из молока (от лат. lactum - молоко). При гидролизе лактоза расщепляется на глюкозу и галактозу: С12Н22О11 + Н2О = С6Н12О6 + С6Н12О6) Сахароза (тростниковый или свекловичный сахара). Название возникло в связи с ее получением либо из сахарной свеклы, либо из сахарного тростника. Тростниковый сахар был известен за много столетий до нашей эры. Лишь в середине XVIII в. этот дисахарид был обнаружен в сахарной свекле и только в начале XIX в. он был получен в производственных условиях. Сахароза очень распространена в растительном мире. Листья и семена всегда содержат небольшое количество сахарозы. Она содержится также в плодах (абрикосах, персиках, грушах, ананасах). Ее много в кленовом и пальмовом соках, кукурузе. Это наиболее известный и широко применяемый сахар. При гидролизе из него образуются глюкоза и фруктоза: С12Н22О11 + Н2О = С6Н12О6 + С6Н12О6.
20. .Биосинтез и ферментативный распад гликогена.
Гликоген - основная форма депонирования глюкозы в клетках животных. У растений эту же функцию выполняет крахмал. В структурном отношении гликоген, как и крахмал, представляет собой разветвленный полимер из глюкозы.Однако гликоген более разветвлен и компактен. Ветвление обеспечивает быстрое освобождение при распаде гликогена большого количества концевых мономеров. Синтез и распад гликогена не являются обращением друг в друга, эти процессы происходят разными путями. Биосинтез гликогена. Гликоген синтезируется в период пищеварения (в течение 1-2 ч после приема углеводной пищи). Гликогенез особенно интенсивно протекает в печени и скелетных мышцах. В начальных реакциях образуется UDF-глюкоза (реакция 3), которая является активированной формой глюкозы, непосредственно включающейся в реакцию полимеризации (реакция 4). Эта последняя реакция катализируется гликогенсинтазой, которая присоединяет глюкозу к олигосахариду или к уже имеющейся в клетке молекуле гликогена, наращивая цепь новыми мономерами. Для подготовки и включения в растущую полисахаридную цепь требуется энергия 1 моль АТР и 1 моль UTP. Ветвление полисахаридной цепи происходит при участии фермента амило - -1,4--1,6-гликозил-трансферазы путем разрыва одной -1,4-связи и переноса олигосахаридного остатка от конца растущей цепи к ее середине с образованием в этом месте -1,6-гликозидной связи. Молекула гликогена содержит до 1 млн остатков глюкозы, следовательно, на синтез расходуется значительное количество энергии. Необходимость превращения глюкозы в гликоген связана с тем, что накопление значительного количества глюкозы в клетке привело бы к повышению осмотического давления, так как глюкоза хорошо растворимое вещество. Напротив, гликоген содержится в клетке в виде гранул, и мало растворим. Распад гликогена - гликогенолиз - происходит в период между приемами пищи.
Распад гликогена. Освобождение глюкозы в виде глюкозо-1-фосфата (реакция 5) происходит в результате фосфоролиза, катализируемого фосфорилазой. Фермент отщепляет концевые остатки один за другим, укорачивая цепи гликогена. Однако этот фермент расщепляет только -1,4 гликозидные связи. Связи в точке ветвления гидролизуются ферментом амило--1,6-гликозидазой, который отщепляет мономер глюкозы в свободном виде.