Уже в составе слюны имеются
 пептидазы типа эндопептидаз, которые могут разрывать внутренние связи в белках и образовывать отдельные полипептидиые цепи — это трипснноподобные ферменты малой активности: саливаин, глан- дулаин, катепсин, калликреин. Считается, что их роль невелика в процессе гидролиза белка, но они повышают эффективность гидролиза пепсином и трипсином.
пептидазы типа эндопептидаз, которые могут разрывать внутренние связи в белках и образовывать отдельные полипептидиые цепи — это трипснноподобные ферменты малой активности: саливаин, глан- дулаин, катепсин, калликреин. Считается, что их роль невелика в процессе гидролиза белка, но они повышают эффективность гидролиза пепсином и трипсином.
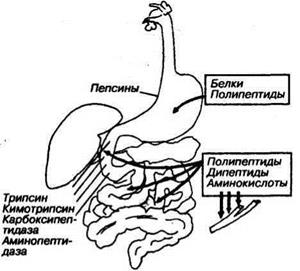
В желудочном соке содержится соляная кислота, создающая высокую концентрацию ионов водорода и вызывающая денатурацию белка, что повышает его гидролиз. HCI также активирует ферменты желудочного сока. В составе желудочного сока содержатся пепсины
Рис. 88. гидролиз веикси. __ — 8 типов. Ферми,™ иырлба-
Кс — кровеносные сосуды. тываются в неактивном виде (пеп-синогены), но под влиянием HCI они активируются. Часть пепсиногенов
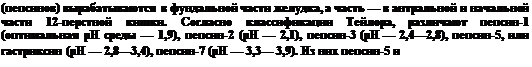 |
пепсин-7, в основном, продуцируются в пилорической части желудка. В основном, пепсины вызывают створаживание молока и разрушают желатиназу. Наличие нескольких типов пепсинов объясняется тем, что в желудке, где длительное время депонируется пища, возникает необходимость набора ферментов, работающих при различных pH, так как концентрация водородных ионов внутри химуса существенно отличается от концентрации этих ионов у стенок желудка.
Итак, в желудке из белков образуются полипептидиые цепочки, которые в дальнейшем под влиянием экзопептидаз (карбоксипептидаз, аминопептидаз) превращаются в дипептиды. Это происходит в 12-перстной кишке и тощей кишке. Сюда изливаются сок панкреатической железы, желчь, желудочный сок.
В панкреатической железе вырабатываются в неактивном виде такие ферменты как трипсиноген, химотрипсиноген, проэластаза, прокарбоксипептидазы А и В, аминопептидаза. Выйдя в полость 12-перстной кишки, они активируются. Процесс начинается с активации трипсиногена. Под влиянием фермента, продуцируемого в кишечнике, — энтерокиназы (энтеропептидазы) трипсиноген переходит в трипсин. В дальнейшем под влиянием трипсина все остальные белки переходят в активную форму (химотрипсиноген —► химотрипсин, прокарбоксипептидаза -» карбоксипептидаза, проэластаза -» эластаза). Итак, трипсин, химотрипсин, эластаза как эндопептидазы расщепляют белки, которые не расщепились пепсинами в желудке, до полипептидов, а карбоксипептидазы А и В, аминопептидаза — до пептидов и дипептидов. В железах тонкого кишечника вырабатываются, главным образом, карбоксипептидазы, аминопептидазы и дипептидазы. Все эти ферменты осуществляют вну- триполостное пищеварение (проводят гидролиз внутри кишки). На апикальной мембране энтероцитов располагаются, согласно данным А. М. Уголева, карбоксипептидазы, аминопептидазы, дипептидазы. В результате их действия образуются аминокислоты, которые тут же передаются на «транспортеры», осуществляющие вторично-активный транспорт, зависимый от ионов натрия. Он осуществляется по такому же механизму, как и транспорт глюкозы (см. выше). Существует как минимум 4 вида аминокислотных «транспортера»: 1) для переноса нейтральных аминокислот (валина, фенилаланина, аланина), 2) для переноса основных аминокислот (аргинина, цистина, лизина, орнитина), 3) для переноса иминокислот (пролина, гидроксипролина) и глицина, 4) для переноса дикарбоновых кислот (глутамимо- вой кислоты, аспарагиновой кислоты). Такое разделение основано на существовании наследственных заболеваний, при которых вследствие генетического дефекта не всасывается группа аминокислот, при этом нарушается и реабсорбция этих же аминокислот в почках. Так, известна болезнь Хартнупа, при которой нарушено всасывание нейтральных аминокислот — валина, фенилаланина, аланина — в тонком кишечнике и в почках, болезнь ими- ноглицинурия — нарушается транспорт пролина, гидроксипролина и глицина, болезнь ци- стинурия — нарушен транспорт цистина, лизина, аргинина и орнитина, синдром Фанкони, при котором имеется аминоацидурия и нарушение всасывания в тонком кишечнике глутаминовой и аспарагиновой аминокислот. Все аминокислоты, а также некоторые белковые молекулы, всосавшиеся путем эндо-экзоцитоза, с током венозной крови через портальную вену доставляются к печени, где они используются для различных целей, в том числе для синтеза белка, для нужд энергетики (например, в процессах глюконеогенеза).
ЖИРЫ, ФОСФОЛИПИДЫ, ХОЛЕСТЕРИН
Жиры, или липиды, представлены в пищевых продуктах в виде триглицеридов (глицерин +3 жирные кислоты), фосфолипидов (глицерин + жирная кислота + фосфорная кислота + аминоспирты), гликолипидов (глицерин + жирная кислота + углеводы), холестерина, стероидов.
Жиры животного происхождения, в основном, содержат насыщенные жирные кислоты, поэтому имеют высокую температуру плавления и в обычном виде они твердые (шпик, сливочное масло, жирная свинина). Жир растительного происхождения, в основном, содержит ненасыщенные жирные кислоты (содержат двойные связи) и имеет низкую температуру плавления — это растительные масла, орехи, овсяная и гречневая крупы. Среди ненасыщенных жирных кислот имеются так называемые незаменимые кислоты — линоленовая и линолевая кислоты, которые не синтезируются в организме, но они необходимы ему, так как из них образуется арахидоновая кислота — предшественница простагландинов и лейкотриенов. Линолевой кислоты много в подсолнечном масле (до 60%). В целом, за сутки необходимо около 80—100 г жиров, из них 30% — растительного происхождения (чтобы получить незаменимые жирные кислоты). Примерно 90% пищевого жира — это триглицериды.
Гидролиз жира происходит, главным образом, с помощью полостного пищеварения с участием липаз и фосфолипаз. Липаза гидролизует жир до жирных кислот и моноглицерида (обычно до 2-моноглицерида).
325
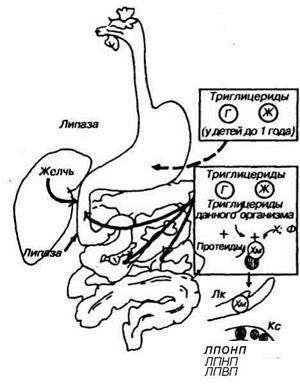 Небольшое количество липазы образуется мелкими слюнными железами корня языка (лингвальная липаза). Железы желудка тоже продуцируют липазу, однако она неактивна в кислой среде. У новорожденных липаза желудочного сока способна расщеплять молочный жир. Ведущую роль в переваривании пищевого жира играет панкреатическая липаза, а также кишечная липаза. Липазы совершают гидролиз в полости кишки, но для эффективного гидролиза поверхность жира должна быть максимальной — это достигается эмульгированием жира с помощью желчных кислот и их солей (холевой, хенодеэок- сихолевой, гликохолевой, таурохс- левой кислотами). Активность липаз возрастает под влиянием так называемой колипазы—фактора, который связывается с липазой и повышает ее способность расщеплять жиры. Ионы кальция тоже повышают активность липазы.
Небольшое количество липазы образуется мелкими слюнными железами корня языка (лингвальная липаза). Железы желудка тоже продуцируют липазу, однако она неактивна в кислой среде. У новорожденных липаза желудочного сока способна расщеплять молочный жир. Ведущую роль в переваривании пищевого жира играет панкреатическая липаза, а также кишечная липаза. Липазы совершают гидролиз в полости кишки, но для эффективного гидролиза поверхность жира должна быть максимальной — это достигается эмульгированием жира с помощью желчных кислот и их солей (холевой, хенодеэок- сихолевой, гликохолевой, таурохс- левой кислотами). Активность липаз возрастает под влиянием так называемой колипазы—фактора, который связывается с липазой и повышает ее способность расщеплять жиры. Ионы кальция тоже повышают активность липазы.
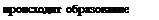 |
 В результате действия липаз образуются мельчайшие капельки, называемые мицеллами (диаметр их примерно 100 нм), которые идут к апикальной мембране энтероцита и в силу своей жирорастворимости входят в энтероцит. Здесь комплекс распадается и происходит синтез триглицеридов, специфичных для человека. Вместе с фос-фолипидами, холестерином и белком-апопротеином AI и В в клетке образуются микровезикулы —так называемые хиломикроны или липопротеин- хиломикроны. Одновременно в клетке образуются и липопротеины очень низкой плотности (тоже в виде микровезикул). Эти структуры проходят через базально-латеральную мембрану энтероцита в интерстиций, а затем переходят в лимфатические сосуды, от них в кровь. Одновременно часть хиломикронов попадает в печень, где синтезируются липопротеины низкой плотности (ЛПНП) и липопротеины высокой плотности (ЛПВП).
В результате действия липаз образуются мельчайшие капельки, называемые мицеллами (диаметр их примерно 100 нм), которые идут к апикальной мембране энтероцита и в силу своей жирорастворимости входят в энтероцит. Здесь комплекс распадается и происходит синтез триглицеридов, специфичных для человека. Вместе с фос-фолипидами, холестерином и белком-апопротеином AI и В в клетке образуются микровезикулы —так называемые хиломикроны или липопротеин- хиломикроны. Одновременно в клетке образуются и липопротеины очень низкой плотности (тоже в виде микровезикул). Эти структуры проходят через базально-латеральную мембрану энтероцита в интерстиций, а затем переходят в лимфатические сосуды, от них в кровь. Одновременно часть хиломикронов попадает в печень, где синтезируются липопротеины низкой плотности (ЛПНП) и липопротеины высокой плотности (ЛПВП).
 ческих и пластических процессов.
ческих и пластических процессов.
ВОДА
В ЖКТ за сутки с пищей и питьем поступает около 2—2,5 л воды. Кроме того, в ЖКТ вода выделяется в составе соков — объемом до 6—7 литров. Итого, в полость ЖКТ поступает до 9,5 л/сутки. Небольшое количество этой воды всасывается в желудке, большая часть — в тонком и толстом кишечнике. В большинстве случаев всасывание воды осуществляется за счет осмоса: реабсорбция осмотически активных веществ типа ионов натрия, кальция, магния, калия, веществ типа глюкозы, аминокислот вызывает пассивную реабсорбцию воды. За счет механизма персорбции часть воды покидает полость кишечника через межэпителиальные щели (путем фильтрации в интерстиций), где с участием поворотно-противоточного механизма создается гипертоническая среда, вызывающая движение воды в интерстиций.
НАТРИЙ
Суточная потребность в натрии составляет около 2—3 г. Его концентрация в крови в среднем 140 ммоль/л. Всасывание натрия осуществляется различными способами: пассивно по градиенту концентрации, активно за счет работы натриевых и натрий-калиевых насосов (в тонком кишечнике, преимущественно, натриевый насос, в толстом — натрий-калиевый насос), и путем персорбции через межэпителиальные поры. Всасывание натрия играет исключительно важную роль, так как от него зависит транспорт аминокислот, глюкозы и других веществ.
На процессы всасывания натрия влияют минералокортикоиды (за счет повышения синтеза натриевых насосов) и гормоны ЖКТ, которые угнетают реабсорбцию натрия в кишечнике: гастрин, секретин, холецистокинин-панкреозимин.
КАЛИЙ
Суточная потребность составляет 2—3 г., столько же, сколько натрия. Концентрация калия в крови в среднем 4,5 ммоль/л. Всасывание осуществляется в тонком и толстом кишечнике за счет пассивного транспорта и с помощью калий-натриевого насоса.
КАЛЬЦИЙ •*■
Суточная потребность кальция составляет 0,7—0,8 г. Концентрация его в плазме крови составляет в среднем 2,25—2,50 ммоль/л. Всасывание кальция происходит весьма медленно и требует наличия специального кальциевого переносчика («транспортера») — кальцийтранспортирующего белка. Его синтез контролируется рядом факторов, в том числе метаболитом витамина Дз. Витамин Дз образуется в коже под влиянием ультрафиолетового облучения из 7-дигидрохолестерина (витамин Дз — холекальциферол). Для того, чтобы он оказывал свой эффект, этот витамин (прогормон) должен пройти две стадии конверсии: в печени под влиянием фермента 25-гидроксилазы он превращается в 25-гидроксихолекаль- циферол (25—ОН Дз), а затем в почках также путем гидроксилирования под влиянием фермента 1-гидроксилазы он превращается в 1,25-дигидроксихолскальциферол (1,25—(ОН)аДэ). Это соединение (гормон) активирует синтез кальцийтранспортирующего белка. При отсутствии этого гормона транспорт кальция нарушается. Гормон паращитовидной железы — паратгормон — способствует конверсии витамина Дз в печени и тем самым повышает эффективность всасывания кальция, что сказывается на уровне кальция в крови.
В растениях содержится витамин Дг — эргокальциферол, в организме он проходит те же стадии конверсии и превращается в 1,25-(ОН)гД2 — по биологической активности он эквивалентен 1,25-(ОН)гДз.
МАГНИЙ
Суточная потребность в магнии — 0,22—0,26 г. Концентрация в крови — около 0,5 ммоль/л. Транспортируется тем же механизмом, что и ионы кальция. Регуляция — аналогична.
ЖЕЛЕЗО
В организме содержится около 3—6 г железа. Из них 800 мг способно мобилизоваться, т. е. быть использованным для синтеза железосодержащих структур, например, гемоглоби-
на. Из 3—6 г железа 65—70% находится в составе гемоглобина эритроцитов, около 20% в мышцах — в составе миоглобина, 10— 15% — в печени и селезенке и около 1% — в составе геминовых ферментов и белков, содержащих негеминовое железо. В среднем за сутки в результате разрушения эритроцитов высвобождается 26 мг железа, из которых 25 мг вновь поступает на синтез гема, а 1 мг выводится, в основном, с желчью. Таким образом, необходимо потребление 1 мг железа в сутки. В тонком кишечнике, где всасывается железо, из пиши извлекается только 10% железа, поэтому суточная потребность с пищей в железе составляет 10—20 мг (при менструациях потребность возрастает в 1,5—2 раза). Учитывая, что диеты в Европе содержат мало железа (на грани минимальной суточной потребности), часто при потере крови возникает дефицит железа, что проявляется, например, в повышенной утомляемости, в снижении работоспособности, головными болями, а при хронической недостаточности — железодефицитной анемией.
В энтероцитах тонкого кишечника, где совершается всасывание железа, имеется «транспортер», который за счет механизмов активного транспорта переносит железо через апикальную мембрану энтероцита внутрь клетки. Здесь двухвалентное железо (а из пиши всасывается в основном двухвалентное железо, поэтому в пище есть восстановители, которые переводят трехвалентное железо в двухвалентное) связывается специфическим белком, который переносит железо в кровь. В крови имеется специальный белок — трансферрин, который транспортирует железо к месту его использования (железо в составе трансферрина находится в форме трехвалентного). В тканях — потребителях железа (печень, селезенка, костный мозг) имеется специальный белок — апоферритин, который, подобно гемоглобину, способен насыщаться двухвалентным железом и превращаться в ферритин, т. е. белок, депонирующий железо. На 1 молекулу этого белка может приходиться до 2500 атомов железа.
Итак, в процессе всасывания и транспорта железа к тканям-потребителям железо меняет валентность с 2-хна 3-х, а затем — вновь на 2-х. Это, вероятно, катализируется специальными окислительно-восстановительными ферментами или самыми белками-переносчиками.
ЙОД, ФТОР
Потребность в йоде — 150 мг/сутки. При недостатке — развитие эндемического зоба (гипотиреоз). Потребность во фторе 1 мг в сутки. Передозировка наступает при поступлении 5 мгв сутки, она вызывает интоксикацию.
ФОСФОР
Суточная потребность в нем составляет 0,7—0,8 г. Концентрация в плазме — около 1 ммоль/л. Метаболиты витамина Д, или Д2 регулируют его всасывание в тонком кишечнике. Паратгормон обычно вызывает снижение уровня фосфора в крови, а кальцитонин, наоборот, повышает. По-видимому, система всасывания фосфора и кальция между собой сопряжена.
ХЛОР
Потребность в нем составляет около 3—5 г в сутки. Концентрация в крови — около 100 ммоль/л. При этом 90% хлора находится во внеклеточной жидкости. Механизм всасывания в основном пассивный — за счет электрохимического градиента, т. е. ионы хлора всасываются вслед за ионами натрия, кальция, калия.
ВИТАМИНЫ
Водорастворимые витамины всасываются в дистальном отделе тощей кишки и проксимальном отделе подвздошной кишки, а жирорастворимые витамины — в средней части тощей кишки. При этом для жирорастворимых витаминов важно наличие желчных кислот.