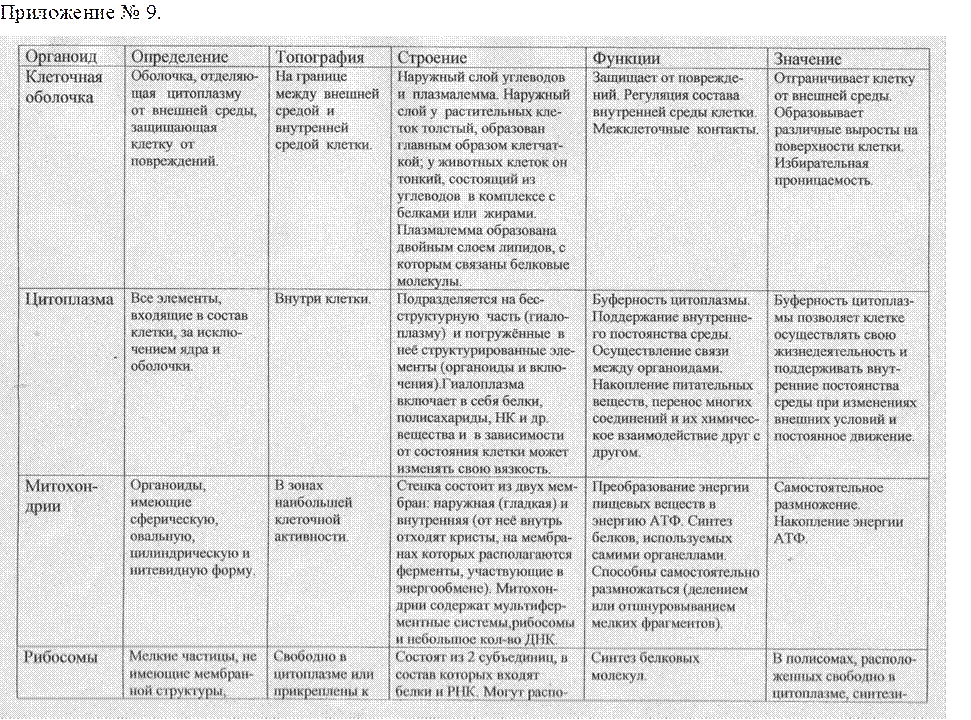
Cходство и различия в строении клеток растений и животных.
Сходство в строении клеток эукариот.
Сейчас нельзя с полной уверенностью сказать, когда и как возникла на Земле жизнь. Мы также точно не знаем, как питались первые живые существа на Земле: автотрофно или гетеротрофно. Но в настоящее время на нашей планете мирно сосуществуют представители нескольких царств живых существ. Несмотря на большое различие в строении и образе жизни, очевидно, что между ними сходств больше, чем различий, и все они, вероятно, имеют общих предков, живших в далёкой архейской эре. О наличии общих «дедушек» и «бабушек» свидетельствует целый ряд общих признаков у клеток эукариот: простейших, растений, грибов и животных. К этим признакам можно отнести:
-общий план строения клетки: наличие клеточной мембраны, цитоплазмы, ядра, органоидов;
-принципиальное сходство процессов обмена веществ и энергии в клетке;
-кодирование наследственной информации при помощи нуклеиновых кислот;
-единство химического состава клеток;
-сходные процессы деления клеток.
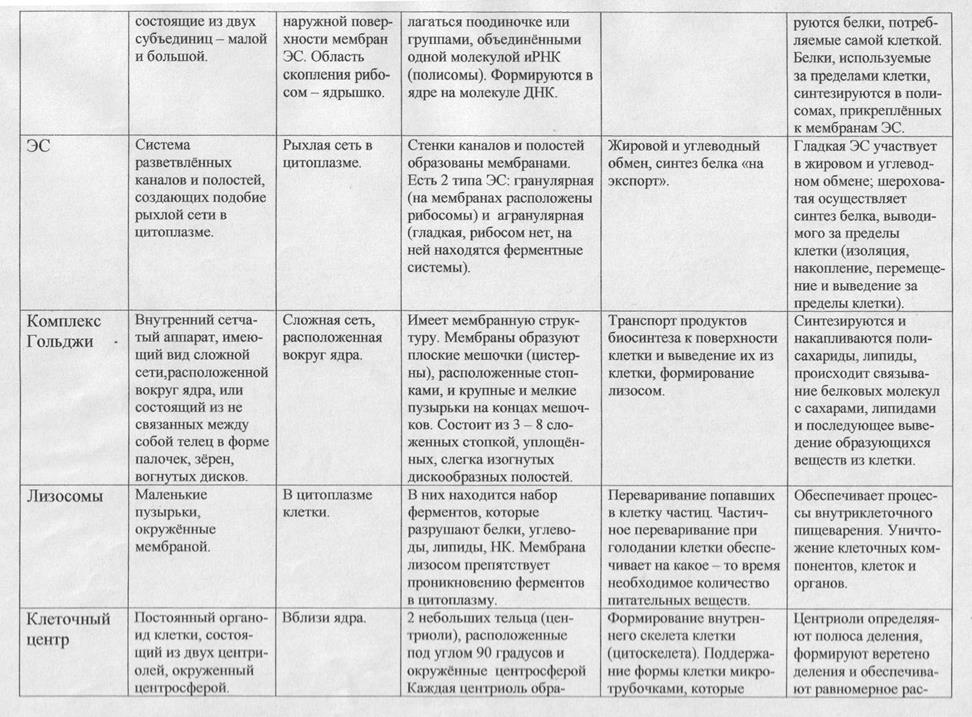
Различия в строении клеток растений и животных.
В процессе эволюции, в связи с неодинаковыми условиями существования клеток представителей различных царств живых существ, возникло множество отличий. Сравним строение и жизнедеятельность клеток растений и животных.
Главное отличие между клетками этих двух царств заключается в способе их питания. Клетки растений, содержащие хлоропласты, являются автотрофами, т. е. Источником углерода для синтеза собственных органических веществ для них являются органические вещества, поступающие с пищей. Эти же пищевые вещества, например углеводы, служат для животных источником энергии. Есть и исключения, такие как зеленые жгутиконосцы, которые на свету способны к фотосинтезу, а в темноте питаются готовыми органическими веществами. Для обеспечения фотосинтеза в клетках растений содержатся пластиды, несущие хлорофилл и другие пигменты.
Так как растительная клетка имеет клеточную стенку, защищающую её содержимое и обеспечивающую постоянную её форму, то при делении между дочерними клетками образуется перегородка, а животная клетка, не имеющая такой стенки, делится с образованием перетяжки.
Сравнение клеток растений и животных
| Признаки | Клетки растений | Клетки животных |
| Способ питания | Автотрофы | Гетеротрофы |
| Клеточная стенка | Есть. Клетка не меняет своей формы. | Клетка может менять свою форму. |
| Пластиды | Хлоропласты, хромопласты, лейкопласты. | Нет. |
| Вакуоли | Немногочисленные крупные полости, заполненные клеточным соком. Содержат запас питательных веществ. Обеспечивают тургорное давление. | Многочисленные мелкие пищеварительные, у некоторых – сократительные. Строение не такое, как у вакуолей растений. |
| Синтез АТФ | В пластидах и митохондриях. | В митохондриях. |
| Запасной углевод | Крахмал. | Гликоген. |
| Способ хранения питательных веществ | Чаще располагаются в клеточном соке вакуоли. | Расположены в цитоплазме в виде клеточных включений. |
| Центриоли | Нет. | Есть. |
| Деление | Образуется перегородка между дочерними клетками. | Образуется перетяжка между дочерними клетками. |
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ, тонкие (не более 10 нм толщиной) липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который включены молекулы разнообразных белков. Расположены на поверхности клеток (плазматическая мембрана) и внутриклеточных частиц (ядра, митохондрий и др.). В весовом отношении в зависимости от типа мембраны на долю липидов приходится 25-60%, а на долю белков — 40-75%. В состав многих мембран входят углеводы, количество которых может достигать 2-10%.
Липиды мембран
Состав липидов биологических мембран очень разнообразен. Характерными представителями липидов клеточных мембран являются фосфолипиды, сфингомиелины и холестерин (стероидный липид). Характерной особенностью мембранных липидов является разделение их молекулы на две функционально различные части: не полярные, не несущие зарядов хвосты, состоящие из жирных кислот, и заряженные полярные головки. Полярные головки несут на себе отрицательные заряды или могут быть нейтральными. Наличие неполярных хвостов объясняет хорошую растворимость липидов в жирах и органических растворителях. В эксперименте, смешивая с водой выделенные из мембран липиды можно получить бимолекулярные слои или мембраны толщиной около 7,5 нм, где периферические зоны слоя — это гидрофильные полярные головки, а центральная зона — незаряженные хвосты молекул липидов. Такое же строение имеют все естественные клеточные мембраны. Клеточные мембраны сильно отличаются друг от друга по составу липидов. Например, плазматические мембраны клеток животных богаты холестерином (до 30%), и в них мало лецитина, в то время как мембраны митохондрий богаты фосфолипидами и бедны холестерином. Липидные молекулы могут перемещаться вдоль липидного слоя, могут вращаться вокруг своей оси, а также переходить из слоя в слой. Белки, плавающие в «липидном озере», тоже обладают некоторой латеральной подвижностью. Состав липидов по обе стороны мембраны различен, что определяет асимметричность в строении билипидного слоя.
Мембранные белки
Белковые молекулы, которые значительно тяжелее и крупнее молекул липидов, как бы вкраплены в билипидный слой мембраны. Большая часть молекул белков частично погружена в мембрану и связана с липидами путем гидрофобных взаимодействий. Это так называемые интегральные белки. Другие белки связаны только с полярными «концами» молекул липидов и находятся на поверхности билипидного слоя, третьи пронизывают мембрану насквозь. Асимметрия в строении мембран ярче всего проявляется в отношении интегральных белков. Они строго ориентированы: их N-концы смотрят в полость вакуолей или во внеклеточную среду.
Мембраны сильно различаются по составу белков, которые собственно и определяют их функциональные свойства. Все мембранные белки можно разделить на три группы: ферменты, рецепторные белки и структурные белки.
В разных мембранах существует характерный набор ферментов. Например, в плазматической мембране локализуется К-Nа-зависимая АТФ-аза, участвующая в транспорте ионов. Рецепторные белки специфически связываются с теми или иными веществами и как бы «узнают» их: это белки-рецепторы для гормонов, для узнавания поверхности соседних клеток, вирусов и т. д. Структурные белки обеспечивают прочность мембраны и связаны с разнообразными белковыми структурами цитоплазмы. Например, в эпителиальных клетках специальные белки плазматической мембраны связываются с элементами цитоскелета и участвуют в образовании целого ряда межклеточных соединений, таких как десмосомы, адгезивные контакты и др.
Углеводы мембран
С мембранами связаны главным образом гликопротеины — молекулы белков, ковалентно связанных с цепочками углеводов. Как правило, молекулы углеводов расположены в наружных слоях мембран. Чаще всего это короткие линейные или разветвленные цепочки, в состав которых входят галактоза, манноза, фруктоза, сахароза, N-ацетилглюкозамин, арабиноза, ксилоза и др.
Синтез клеточных мембран
Строятся все клеточные мембраны (кроме мембран митохондрий и пластид) в гранулярном эндоплазматическом ретикулюме. От него отщепляются мелкие мембранные вакуоли, которые сливаются с мембранами аппарата Гольджи. Мембранные везикулы, продуцируемые, в свою очередь, аппаратом Гольджи, идут на построение плазматической мембраны, секреторных вакуолей и всех других мембранных компонентов клетки.
ПЛАЗМАТ?ИЧЕСКАЯ МЕМБР?АНА (плазмалемма, клеточная мембрана), поверхностная, периферическая структура, окружающая протоплазму растительных и животных клеток. Служит не только механическим барьером, но, главное, ограничивает свободный двусторонний поток в клетку и из нее низко- и высокомолекулярных веществ. Более того, плазмалемма выступает как структура, «узнающая» различные химические вещества и регулирующая избирательный транспорт этих веществ в клетку. Как и другие мембраны клетки, она возникает и обновляется за счет синтетической активности эндоплазматического ретикулюма и имеет сходное с ними строение.
Барьерно-транспортная роль плазмалеммы.
Механическая устойчивость плазматической мембраны определяется не только свойствами самой мембраны, но и свойствами прилежащих к ней гликокаликса и кортикального слоя цитоплазмы.
Внешняя поверхность плазматической мембраны покрыта рыхлым волокнистым слоем вещества толщиной 3-4 нм — гликокаликсом. Он состоит из ветвящихся полисахаридных цепей мембранных интегральных белков, между которыми могут располагаться выделенные клеткой гликолипиды и протеогликаны. Тут же обнаруживаются некоторые клеточные гидролитические ферменты, участвующие во внеклеточном расщеплении веществ (внеклеточное пищеварение, например, в эпителии кишечника). Кортикальный слой цитоплазмы, толщиной 0,1-0,5 мкм, не содержит рибосом и мембранных структур, но богат актиновыми микрофиламентами.
Плазматическая мембрана, как и другие липопротеидные мембраны клетки, является полупроницаемой. Максимальной проникающей способностью обладает вода и растворенные в ней газы. Транспорт ионов может проходить по градиенту концентраций, т. е. пассивно, без затрат энергии. В этом случае некоторые мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые ионы проходят сквозь мембрану за счет простой диффузии. В других случаях специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану. Такой тип переноса называется активным транспортом и осуществляется с помощью белковых ионных насосов. Например, затрачивая 1 молекулу АТФ, система К-Nа насоса откачивает за один цикл из клетки 3 иона Nа и закачивает 2 иона К против градиента концентрации. В сочетании с активным транспортом ионов через плазмалемму проникают различные сахара, нуклеотиды и аминокислоты. Макромолекулы, такие как, например, белки, через мембрану не проходят. Они, а также более крупные частицы вещества транспортируются внутрь клетки посредством эндоцитоза. При эндоцитозе определенный участок плазмалеммы захватывает, обволакивает внеклеточный материал, заключает его в мембранную вакуоль. Эта вакуоль — эндосома — сливается в цитоплазме с первичной лизосомой и происходит переваривание захваченного материала. Эндоцитоз формально разделяют на фагоцитоз (поглощение клеткой крупных частиц) и пиноцитоз (поглощение растворов). Плазматическая мембрана принимает участие и в выведении веществ из клетки с помощью экзоцитоза — процесса, обратного эндоцитозу.
Рецепторная роль плазмалеммы
Белки-переносчики внешней мембраны клетки являются также рецепторами, узнающими определенные ионы и взаимодействующими с ними. В качестве рецепторов на поверхности клетки могут выступать белки мембраны или элементы гликокаликса. Такие чувствительные к отдельным веществам участки разбросаны по поверхности клетки или собраны в небольшие зоны. Роль многих клеточных рецепторов заключается не только в связывании специфических веществ, но и в передаче сигналов с поверхности внутрь клетки. Например, при действии гормона на клетку цепь событий развертывается следующим образом: молекула гормона специфически взаимодействует с рецепторным белком плазмалеммы и, не проникая в клетку, активирует фермент, синтезирующий ЦАМФ. Последний активирует или ингибирует внутриклеточный фермент или группу ферментов.
Разнообразие и специфичность наборов рецепторов на поверхности клеток приводит к созданию очень сложной системы маркеров, позволяющих клеткам отличать «своих» (той же особи или того же вида) от «чужих».
Межклеточные соединения
У многоклеточных организмов за счет межклеточных взаимодействий образуются сложные клеточные ансамбли. При тесном соседстве клеток друг с другом гликокаликс обеспечивает слипание клеток за счет присутствия в нем трансмембранных гликопротеидов кадгеринов. Это простой межклеточный контакт, при котором зазор между клетками составляет 10-20 нм. В эпителиях часто встречается плотное, или запирающее, соединение, при котором внешние слои двух плазматических мембран максимально сближены и в точках их соприкосновения лежат глобулы интегральных белков мембраны. Такой контакт непроницаем для молекул и ионов, он запирает межклеточные полости.
Заякоривающие соединения, или контакты, не только соединяют плазматические мембраны соседних клеток, но и связываются с фибриллярными элементами цитоскелета. Например, для десмосом, имеющих вид бляшек или кнопок, в межклеточном пространстве характерно наличие плотного слоя гликопротеидов десмоглеинов. С цитоплазматической стороны к плазмалемме прилежит слой белка десмоплакина, связанный с промежуточными филаментами цитоскелета.
Щелевые контакты считаются коммуникационными соединениями клеток. В зоне щелевого контакта может быть от 20-30 до нескольких тысяч коннексонов — цилидрических белковых структур с внутренним каналом диаметром 2 нм. Каждый коннексон состоит из 6 субъединиц белка коннектина. Коннексоны играют роль прямых межклеточных каналов, по которым ионы и низкомолекулярные вещества могут диффундировать из клетки в клетку.
ЦИТОПЛАЗМА (от цито... и плазма), внеядерная часть протоплазмы животных и растительных клеток. Состоит из гиалоплазмы, в которой содержатся органоиды и др. включения.
* * *
ЦИТОПЛАЗМА (от цито... и плазма), внеядерная часть протоплазмы клетки, то есть внутреннее содержимое клетки без ядра. Термин предложен Э. Страсбургером (1882).
Экспериментально можно получить живые безъядерные клетки-цитопласты, которые в течение 1-3 суток могут синтезировать белки, липиды, АТФ. Затем они, конечно, погибают из-за невозможности синтеза новых РНК в отсутствии ядра. Объем цитоплазмы у разных клеток неодинаков: в лимфоцитах он примерно равен объему ядра, а в клетках печени цитоплазма составляет 94% общего объема клетки. Формально в цитоплазме различают три части: органеллы, включения и гиалоплазма. Органеллы — обязательные для любой клетки компоненты, без которых клетка не может поддерживать свое существование. К одномембранным относятся органеллы вакуолярной системы: эндоплазматический ретикулюм, лизосомы, аппарат Гольджи, пероксисомы, а также плазматическая мембрана. Двухмембранные органеллы — это митохондрии и пластиды. К этой группе можно отнести и клеточное ядро.
Общим свойством мембранных органелл является то, что они построены из липопротеидных мембран, замыкающихся сами на себя, так, что образуют замкнутые полости и тем самым разделяют цитоплазму на группы функционально различных отсеков.
К немембранным органеллам принадлежат рибосомы, центросомы животных клеток.
Включения встречаются не всегда и представляют собой отложения запасных веществ (гликоген, желток) или скопления продуктов метаболизма (пигменты, кристаллы солей в растениях).
Гиалоплазма (от «hyalinе» — прозрачный) — это основная плазма, цитозоль, истинная внутренняя среда клетки. Состав гиалоплазмы весьма сложен, а консистенция приближается к гелю. Гели — структурированные коллоидные системы с жидкой дисперсной средой, которые под воздействием внешних или внутренних факторов могут менять свое агрегатное состояние и переходить в более жидкую фазу — золь. Подобные гель-золь переходы могут происходить в цитоплазме под влиянием белка актина, причем меняется ее состояние в различных участках клетки, что и обеспечивает движение всей клетки. При взаимодействии фибриллярного актина с белками типа фимбрина происходит стабилизация геля, а при связывании с белками, активность которых зависит от концентрации ионов Са (например, гельзолин), вся система переходит в жидкое состояние. Вообще выраженность элементов клеточного скелета, и актинового его компонента в том числе, может значительно меняться в течение клеточного цикла.
Функциональное значение цитоплазмы очень велико. В гиалоплазме, кроме различных ионов неорганических соединений, содержатся ферменты, участвующие в синтезе аминокислот, нуклеотидов, жирных кислот, сахаров. На рибосомах и полирибосомах, сидящих на мембранах, синтезируются разнообразные белки, обеспечивающие клеточный метаболизм.
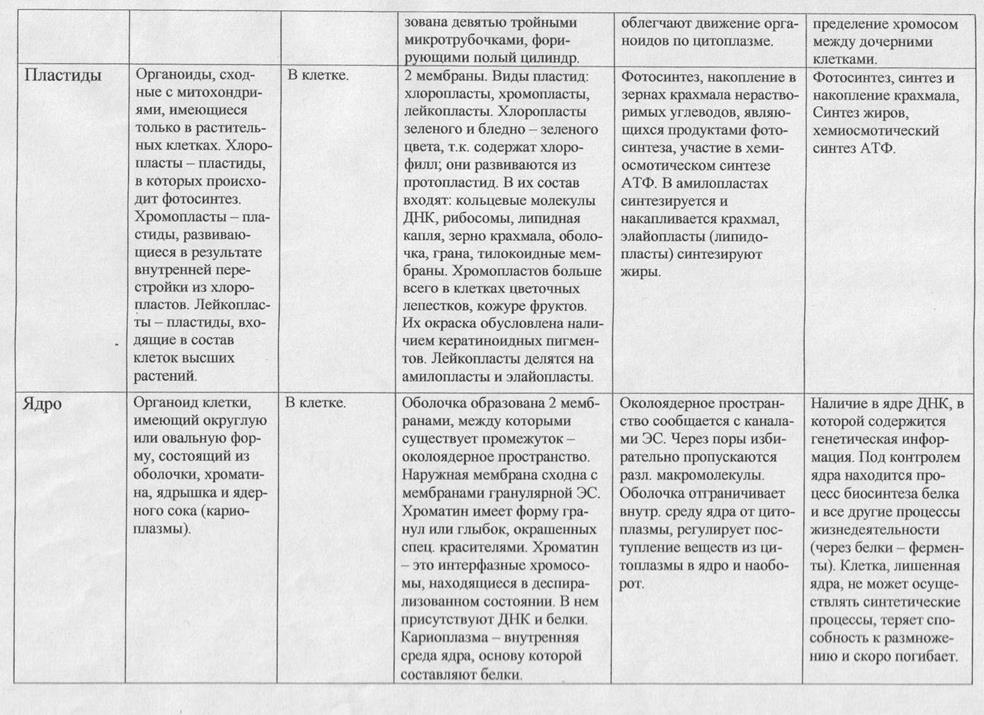
ЯДР?О?, в биологии — обязательная часть клетки у многих одноклеточных и всех многоклеточных организмов. Типичное ядро отделено от окружающей цитоплазмы оболочкой, содержит ядрышко, хромосомы и кариоплазму. Размеры от 1 мкм (у некоторых простейших) до 1 мм (в яйцах некоторых рыб и земноводных). Управляет синтезом белков (в т. ч. ферментов) и через них всеми физиологическими процессами в клетке. Большинство клеток содержит одно ядро. По наличию или отсутствию в клетках оформленного ядра все организмы делят соответственно на эукариот и прокариот.
* * *
ЯДР?О, клеточное ядро, обязательная часть клетки у многих одноклеточных и всех многоклеточных организмов. Все организмы нашей биосферы как одноклеточные, так и многоклеточные, подразделяются на эукариот — их клетки содержат ядро, и прокариот, клетки которых не имеют морфологически оформленного ядра. Термин «ядро» (лат. nucleus) впервые применил Р. Броун в 1833 году, когда описывал шарообразные структуры, наблюдаемые им в клетках растений.
Ядерная оболочка
Внутреннее пространство клеточного ядра отделено от цитоплазмы ядерной оболочкой, состоящей из двух мембран. Мембраны оболочки ядра сходны по строению с другими мембранными компонентами клетки и построены по тому же принципу: это тонкие липопротеидные пленки, состоящие из двойного слоя липидных молекул, в который встроены молекулы белков. Пространство между внутренней и внешней ядерными мембранами называется перинуклеарным. На поверхности внешней ядерной мембраны обычно располагается большое количество рибосом, и иногда удается наблюдать непосредственный переход этой мембраны в систему каналов гранулярной эндоплазматической сети клетки. Внутренняя ядерная мембрана связана с тонким волокнистым белковым слоем — ядерной ламиной, состоящей из белков ламинов. Густая сеть фибрилл ядерной ламины способна обеспечить целостность ядра, даже после растворения липидных мембран оболочки ядра в эксперименте. С внутренней стороны к ламине крепятся петли хроматина, заполняющего ядро.
Ядерная оболочка имеет отверстия диаметром около 90 нм, образующиеся засчет слияния внешней и внутренней ядерных мембран. Такие отверстия в оболочке ядра окружены сложными белковыми структурами, получившими название комплекса ядерной поры. Восемь белковых субъединиц, входящих в состав ядерной поры, располагаются вокруг перфорации ядерной оболочки в виде колец, диаметром около120 нм, наблюдаемых в электронный микроскоп с обеих сторон ядерной оболочки. Белковые субъединицы комплекса поры имеют выросты, направленные к центру поры, где иногда видна «центральная гранула» диаметром 10-40 нм. Размер ядерных пор и их структура стандартны для всех клеток эукариот. Число ядерных пор зависит от метаболической активности клеток: чем выше уровень синтетических процессов в клетке, тем больше пор на единицу площади поверхности клеточного ядра. В процессе ядерно-цитоплазматического транспорта ядерные поры функционируют как некое молекулярное сито, пропуская ионы и мелкие молекулы (сахара, нуклеотиды, АТФ и др.) пассивно, по градиенту концентрации, и осуществляя активный избирательный транспорт крупных молекул белков и рибонуклеопротеидов, то есть комплексов рибонуклеиновых кислот (РНК) с белками. Так, например, белки, транспортируемые в ядро из цитоплазмы, где они синтезируются, должны иметь определенные последовательности примерно из 50 аминокислот, (т. наз. NLS последовательности), «узнаваемые» комплексом ядерной поры. В этом случае комплекс ядерной поры, затрачивая энергию в виде АТФ, активно транслоцирует белок из цитоплазмы в ядро.
Хроматин
Клеточное ядро является вместилищем практически всей генетической информации клетки, поэтому основное содержимое клеточного ядра — это хроматин: комплекс дезоксирибонуклеиновой кислоты (ДНК) и различных белков. В ядре и, особенно, в митотических хромосомах, ДНК хроматина многократно свернута, упакована особым образом для достижения высокой степени компактизации. Ведь все длинные нити ДНК, общая длина которых составляет, например, у человека около 164 см, необходимо уложить в клеточное ядро, диаметр которого всего несколько микрометров. Эта задача решается последовательной упаковкой ДНК в хроматине с помощью специальных белков. Основная масса белков хроматина — это белки гистоны, входящие в состав глобулярных субъединиц хроматина, называемых нуклеосомами. Всего существует 5 видов белков гистонов. Нуклеосома представляет собой цилиндрическую частицу, состоящую из 8 молекул гистонов, диаметром около 10 нм, на которую «намотано» чуть менее двух витков нити молекулы ДНК. В электронном микроскопе такой искусственно деконденсированный хроматин выглядит как «бусины на нитке». В живом ядре клетки нуклеосомы плотно объединены между собой с помощью еще одного линкерного гистонового белка, образуя так называемую элементарную хроматиновую фибриллу, диаметром 30 нм. Другие белки, негистоновой природы, входящие в состав хроматина обеспечивают дальнейшую компактизацию, т. е. укладку, фибрилл хроматина, которая достигает своих максимальнах значений при делении клетки в митотических или мейотических хромосомах. В ядре клетки хроматин присутствует как в виде плотного конденсированного хроматина, в котором 30 нм элементарные фибриллы упакованы плотно, так и в виде гомогенного диффузного хроматина. Количественное соотношение этих двух видов хроматина зависит от характера метаболической активности клетки, степени ее дифференцированности. Так, например, ядра эритроцитов птиц, в которых не происходит активных процессов репликации и транскрипции, содержат практически только плотный конденсированный хроматин. Некоторая часть хроматина сохраняет свое компактное, конденсированное состояние в течение всего клеточного цикла — такой хроматин называется гетерохроматином и отличается от эухроматина рядом свойств.
Репликация и транскрипция
Клетки эукариот содержат обычно несколько хромосом (от двух до нескольких сотен), которые теряют в ядре (в интерфазе, т. е. между митотическоми делениями) клетки свою компактную форму, разрыхляются и заполняют объем ядра в виде хроматина. Несмотря на деконденсированное состояние, каждая хромосома занимает в ядре строго определенное положение и связана с ядерной оболочкой посредством ламины. Строго закреплены на внутренней поверхности оболочки ядра такие структуры хромосом, как центромеры и теломеры. На определенной стадии жизненного цикла клетки, в синтетическом периоде, происходит репликация, т. е. удвоение всей ДНК ядра, и хроматина становится в два раза больше. Белки, необходимые для этого процесса, поступают, конечно, из цитоплазмы через ядерные поры. Таким образом, клетка готовится к предстоящему клеточному делению — митозу, когда общее количество ДНК в ядре вернется к первоначальному уровню.
Реализация генетической информации, заключенной в ДНК в виде генов, начинается с транскрипции, т. е. с синтеза информационных РНК (и-РНК) — точных копий генов, по которым затем будут строиться в цитоплазме на рибосомах белки. Этот процесс проходит в различных точках в обьеме ядра, морфологически ничем не отличающихся от окружающего хроматина. Чаще всего удается наблюдать транскрипцию диффузного, т.е. деконденсированного хроматина.
Кроме хроматина, составляющего хромосомы, в ядрах эукариот обычно содержится одно или несколько ядрышек. Это плотные структуры, не имеющие собственной оболочки и представляющие собой скопления молекул другого типа РНК — рибосомной РНК (р-РНК) в комплексе с белками. Такие комплексы называют рибонуклеопротеидами (РНП). Ядрышки имеют стандартную морфологию и образуются в ядре после деления клетки вокруг постояннодействующих точек активного синтеза рибосомной РНК. Гены рибосомной РНК, в отличие от большинства других генов, кодирующих белки, содержатся в геноме в виде многочисленных копий. Эти копии, расположенные в молекуле ДНК тандемно, т. е. друг за другом, располагаются в определенных районах нескольких хромосом генома. Такие районы хромосом называют ядрышковыми организаторами. Морфологически в ядрышке с помощью электронного микроскопа можно выделить следующие 3 зоны: гомогенные компактные фибриллярные центры, содержащие ДНК ядрышковых организаторов; плотный фибриллярный компонент вокруг них, где идет транскрипция генов рибосомной РНК и массивный гранулярный компонент ядрышка, состоящий из частиц РНП — будущих рибосом. Эти гранулы РНП, образующиеся в ядрышке, транспортируются в цитоплазму и образуют рибосомы, осуществляющие синтез всех белков клетки. Третий основной тип клеточных РНК — мелкие транспортные РНК — транскрибируются в различных участках ядра и выходят в цитоплазму через ядерные поры. Там они, как известно, обеспечивают транспортировку аминокислот к рибосомам в процессе синтеза белков.
Ядерный белковый матрикс
Для осуществления процессов репликации, транскрипции, а также поддержания определенного положения хромосом в обьеме ядра существуют каркасные белковые структуры, называемае ядерным белковым матриксом. Такой матрикс состоит, по крайней мере из трех морфологических компонентов: периферического фиброзного слоя- ламины; внутреннего, или интерхроматинового матрикса ядра и матрикса ядрышка. Наблюдения показывают, что компоненты ядерного матрикса — это не жесткие застывшие структуры, они динамичны и могут сильно видоизменяться в зависимости от функциональных особенностей ядер. Показано, что белковый матрикс имеет множество точек прочного связывания с ДНК ядра, которая, в свою очередь, имеет специальные последовательности нуклеотидов, необходимые для этого.
Ъ
