Ротовая полость
Со слюной сюда поступает кальций-содержащий фермент α-амилаза. Оптимум ее рН 7,1-7,2, активируется ионами Cl–. Являясь эндоамилазой, она беспорядочно расщепляет внутренние α1,4-гликозидные связи и не влияет на другие типы связей.
В ротовой полости углеводы расщепляются до декстринов и мальтозы. Дисахариды не гидролизуются.
Желудок
Из-за низкой рН амилаза инактивируется, хотя некоторое время расщепление углеводов продолжается внутри пищевого комка.
Кишечник
В полости тонкого кишечника работают совместно панкреатическая α-амилаза, разрывающая внутренние α1,4-связи, изомальтаза, разрывающая α1,6-связи изомальтозы, олиго-α1,6-глюкозидаза, действующая на точки ветвления крахмала и гликогена.
Кроме полостного, имеется еще и пристеночное пищеварение, которое осуществляют:
· сахаразо-изомальтазный комплекс (рабочее название сахараза) – в тощей кишке гидролизует α1,2-, α1,4-, α1,6-гликозидные связи, расщепляет сахарозу, мальтозу, мальтотриозу, изомальтозу,
· гликоамилазный комплекс – находится в нижних отделах тонкого кишечника и расщепляет α1,4-гликозидные связи в олигосахаридах,
· β-гликозидазный комплекс (рабочее название лактаза) – гидролизует β1,4-гликозидные связи между галактозой и глюкозой (лактозу). У детей активность лактазы очень высока уже до рождения и сохраняется на высоком уровне до 5-7 лет, после чего снижается.
Роль целлюлозы в пищеварении
Целлюлоза ферментами человека не переваривается. Но в толстом кишечнике под действием микрофлоры до 75% ее количества гидролизуется с образованием целлобиозы и глюкозы. Глюкоза частично используется самой микрофлорой и окисляется до органических кислот (масляной, молочной), которые стимулируют перистальтику кишечника. Частично глюкоза может всасываться в кровь.
Основная роль целлюлозы для человека:
· стимулирование перистальтики кишечника,
· формирование каловых масс,
· стимуляция желчеотделения,
· абсорбция холестерола и других веществ, что препятствует их всасыванию.
10) Гликогенез
Гликогенез - биохимическая реакция протекающая в основном в печени и мышцах в результате которой глюкоза превращается в гликоген. Это очень важная биологическая реакция так как глюкоза является основным источником энергии в организме, а гликоген это форма хранения глюкозы в клетке на случай энергетических затрат между приемами пищи. Гликогенез это только один из этапов сложного превращения углеводов в организме. В целом это происходит следующим образом: Во время приема пищи углеводы, к которым в первую очередь относится крахмал, а также дисахариды сахароза, лактоза и мальтоза, под действием фермента слюны амилазы расщепляются на менее крупные чем крахмал молекулы. Далее в тонком кишечнике уже другие ферменты (панкреатическая амилаза, сахараза, мальтаза и лактаза) гидролизируют углеводные остатки до моносахаридов, одним из которых является глюкоза. Затем половина всосавшейся глюкозы поступает в печень, а остальная часть транспортируется в остальные ткани. Поступление глюкозы в клетки во многих органах регулируется инсулином, за исключением печени и мозга где скорость диффузии зависит только от концентрации глюкозы. В клетках происходит распад глюкозы - гликолиз. Гликолиз может происходить с участием кислорода (аэробный процесс), тогда в результате образуется две молекулы пирувата, и без участия кислорода (анаэробный процесс), в этом случае образуются две молекулы лактата. В случае аэробного процесса пируват в конечном итоге окисляется до углекислого газа и воды. А анаэробный процесс включает те же реакции что и аэробный, только в конце пируват превращается в лактат. В результате распада глюкозы с участием кислорода образуется 38 молекул АТФ - источника энергии в живых организмах, а после анаэробного гликолиза всего 2 молекулы АТФ. Смысл этого заключается в том, что анаэробный процесс протекает в мышцах в начале интенсивной физической нагрузки когда снабжение кислородом ограничено.
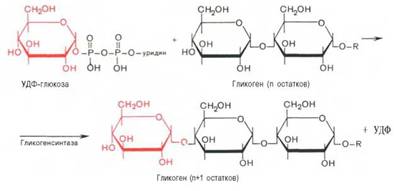
Часть попадающей с пищей глюкозы не идет на образование АТФ, а откладывается про запас в виде гликогена у животных и в виде крахмала у растений. Гликоген и крахмал это разветвленные цепочки из молекул глюкозы. Процесс гликогенеза, т.е. образование гликогена из глюкозы представляет собой процесс полимеризации, т.е происходит последовательное присоединение друг к другу мономеров глюкозы и образование полисахаридной цепи гликогена. Мышечный гликоген является источником глюкозы для самой мышечной клетки, а печеночный поддерживает физиологическую концентрацию глюкозы в крови.
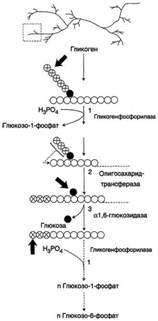
Распад гликогена
Так же как и синтез, расщепление гликогена начинается с нередуцирующего конца полисахаридной цепи. При этом наличие разветвлённой структуры гликогена облегчает быстрое высвобождение глюкозных остатков, так как чем больше концов имеет молекула гликогена, тем больше молекул гликогенфосфорилазы могут действовать одновременно.
Гликогенфосфорилаза расщепляет только α-1,4-гликозидные связи (реакция 1). Последовательное отщепление глюкозных остатков прекращается, когда до точки ветвления остаётся 4 мономера. Подобная особенность в действии гликогенфосфорилазы обусловлена размером и строением её активного центра.
Дальнейший распад гликогена требует участия двух других ферментов. Сначала три оставшихся до точки ветвлении глюкозных остатка переносятся при участии олигосахаридтрансферазы (реакция 2) на нередуцирующий конец соседней цепи, удлиняя её и таким образом создавая условия для действия фосфорилазы. Оставшийся в точке ветвления глюкозный остаток гидролитически отщепляется с помощью α-1,6-глюкозидазы в виде свободной глюкозы (реакция 3), после чего неразветвлённый участок гликогена может вновь атаковаться фосфорилазой.
Считают, что перенос трёх остатков глюкозы и удаление мономера из точки ветвления (реакции 2 и 3) катализирует один и тот же фермент, который обладает двумя разными ферментативными активностями - трансферазной и гликозидазной. Его называют "деветвящим" ферментом (от англ, debranching enzyme).
Продукт действия гликогенфосфорилазы - глюкозо-1-фосфат - затем изомеризуется в глюкозо-6-фосфат фосфоглюкомутазой. Далее глюкозо-6-фосфат включается в процесс катаболизма или другие метаболические пути. В печени (но не в мышцах) глюкозо-6-фосфат может гидролизоваться с образованием глюкозы, которая выделяется в кровь. Эту реакцию катализирует фермент глюкозо-6-фосфатаза. Реакция протекает в просвете ЭР, куда с помощью специального белка транспортируется глюкозо-6-фосфат. Фермент локализован на мембране ЭР таким образом, что его активный центр обращён в просвет ЭР. Продукты гидролиза (глюкоза и неорганический фосфат) возвращаются в цитоплазму также с помощью транспортных систем.
12) гликолиз
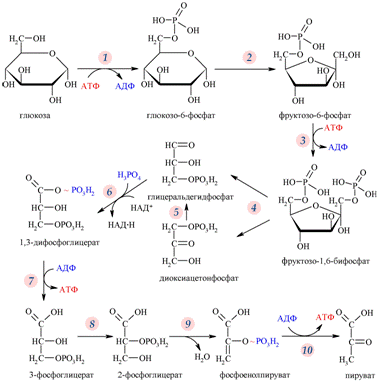
В клетках эукариотических организмов десять ферментов, катализирующих распад глюкозы до ПВК, находятся в цитозоле, все остальные ферменты, имеющие отношение к энергетическому обмену, — вмитохондриях и хлоропластах. Поступление глюкозы в клетку осуществляется двумя путями: натрий-зависимый симпорт (преимущественно для энтероцитов и эпителия почечных канальцев) и облегчённая диффузия глюкозы с помощью белков-переносчиков. Работа этих белков-транспортёров контролируется гормонами и, в первую очередь, инсулином. Сильнее всего инсулин стимулирует транспорт глюкозы вмышцах и жировой ткани. Гликолитический путь представляет собой 10 последовательных реакций, каждая из которых катализируется отдельным ферментом.
Процесс гликолиза условно можно разделить на два этапа. Первый этап, протекающий с расходом энергии 2-х молекул АТФ, заключается в расщеплении молекулы глюкозы на 2 молекулы глицеральдегид-3-фосфата. На втором этапе происходит НАД-зависимое окисление глицеральдегид-3-фосфата, сопровождающееся синтезом АТФ. Сам по себе гликолиз является полностью анаэробным процессом, то есть не требует для протекания реакций присутствия кислорода.
Результатом гликолиза является превращение одной молекулы глюкозы в две молекулы пировиноградной кислоты (ПВК) и образование двух восстановительных эквивалентов в виде кофермента НАД∙H.
Полное уравнение гликолиза имеет вид:
Глюкоза + 2НАД+ + 2АДФ + 2Фн = 2НАД∙Н + 2ПВК + 2АТФ + 2H2O + 2Н+.
При отсутствии или недостатке в клетке кислорода пировиноградная кислота подвергается восстановлению до молочной кислоты, тогда общее уравнение гликолиза будет таким:
Глюкоза + 2АДФ + 2Фн = 2лактат + 2АТФ + 2H2O.
Таким образом, при анаэробном расщеплении одной молекулы глюкозы суммарный чистый выход АТФ составляет две молекулы, полученные в реакциях субстратного фосфорилирования АДФ.
У аэробных организмов конечные продукты гликолиза подвергаются дальнейшим превращениям в биохимических циклах, относящихся к клеточному дыханию. В итоге после полного окисления всех метаболитов одной молекулы глюкозы на последнем этапе клеточного дыхания — окислительном фосфорилировании, происходящем на митохондриальной дыхательной цепи в присутствии кислорода, — дополнительно синтезируются ещё 34 или 36 молекулы АТФ на каждую молекулу глюкозы.
Реакции и ферменты гликолиза
| Последовательность реакции | Реакция гликолиза | Фермент, катализирующий реакцию | Коферменты и кофакторы | Активаторы | Ингибиторы |
| 1. | Глюкоза + АТФ Ûглюкозо-6-фосфат + АДФ | Гексокиназа (глюкокиназа) | Ионы Mg2+ | Комплекс Mg2+ — АТФ4-, Фнеорг. | Глюкозо-6-фосфат, АДФ |
| 2. | Глюкозо-6-фосфат Ûфруктозо-6-фосфат | Фосфоглюкоизомераза | Ионы Mg2+ | Нет | Нет |
| 3. | Фруктозо-6-фосфат + АТФ Ûфруктозо-1,6-ди-фосфат | Фосфофруктокиназа | Ионы Mg2+ | Фнеорг. АМФ, АДФ, циклический 3',5'-АМФ (цАМФ), ионы К+ | Комплекс Mg2+ — АТФ4-, цитрат |
| 4. | Фруктозо-1,6-дифосфат Ûглицеральдегид-3-фосфат + диоксиацетонфосфат | Альдолаза | Нет | Ионы Fe2+, СО2+ | Цистеин, ФФнеорг. |
| 5. | Глицеральдегид-3-фосфат Ûдиоксиацетонфосфат | Фосфотриозоизомераза | Ионы Mg2+ | Нет | Фнеорг. |
| 6. | 2Глицеральдегид-3-фосфат + 2НАД+ + 2Фнеорг. Û2(1,3-дифосфоглицерат) + 2НАДН + 2Н+ | Глицеральдегидфосфат-дегидрогеназа | НАД+ | Арсенат | Йодоацетат |
| 7. | 2(1,3-дифосфоглицерат) + 2АДФ Û 2(3-фосфоглицерат) + 2АТФ | Фосфоглицераткиназа | Ионы Mg2+ | Нет | Не известны |
| 8. | 2 (3-фосфоглицерат)Û 2(2-фосфоглицерат) | Фосфоглицеромутаза | Ионы Mg2+, 2,3-дифосфогли церат | Нет | Не известны |
| 9. | 2(3-фосфоглицерат)Û2фосфоенолпируват | Енолача | Ионы Mg2+, Mn2+ | Нет | Ионы F-, Са2+, Фнеорг. |
| 10. | 2Фосфоенолпируват + 2АДФ Û 2пируват + 2АТФ | Пируваткиназа | Ионы К+, фруктозо-1,6-дифосфат | Нет | С2+, АТФ, аланин, жирные кислоты ацетил-КоА |
| 11. | 2Пируват + 2 НАД×Н + 2Н+ Û 2лактат + 2НАД+ | Лактатдегидрогеназа | НАД+ | Нет | Не известны |
13) окислительное декарбоксилирование
Окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
На I стадии этого процесса пируват (рис. 10.8) теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ) в составеактивного центра фермента пируватдегидрогеназы (E1). На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованиемацетильной группы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментом дигидроли-поилацетилтрансферазой (Е2). Этот фермент катализирует III стадию – перенос ацетильной группы на коэнзим КоА (HS-KoA) с образованием конечного продукта ацетил-КоА, который является высокоэнергетическим (макроэргическим) соединением.
На IV стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамид–Е2. При участии ферментадигидролипоилдегидрогеназы (Е3) осуществляется перенос атомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на ФАД, который выполняет роль простетической группы данного фермента и прочно с ним связан. На V стадии восстановленный ФАДН2 дигидро-липоилдегидрогеназы передает водород на кофермент НАД с образованием НАДН + Н+.
Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа, ди-гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из которых три относительно прочно связаны с ферментами (ТПФ-E1, ли-поамид-Е2 и ФАД-Е3), а два – легко диссоциируют (HS-KoA и НАД).
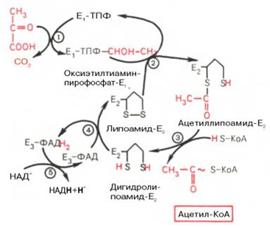
Все эти ферменты, имеющие субъединичное строение, и коферменты организованы в единый комплекс. Поэтому промежуточные продукты способны быстро взаимодействовать друг с другом. Показано, что составляющие комплекс полипептидные цепи субъединиц дигидролипоил-ацетилтрансферазы составляют как бы ядро комплекса, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа. Принято считать, что нативный ферментный комплекс образуется путем самосборки.
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
Пируват + НАД+ + HS-KoA –> Ацетил-КоА + НАДН + Н+ + СO2.
Реакция сопровождается значительным уменьшением стандартной свободной энергии и практически необратима.
Образовавшийся в процессе окислительного декарбоксилирования аце-тил-КоА подвергается дальнейшему окислению с образованием СО2 и Н2О. Полноеокисление ацетил-КоА происходит в цикле трикарбоновых кислот (цикл Кребса). Этот процесс, так же как окислительное декарбо-ксилирование пирувата, происходит в митохондриях клеток.
14)цикл пентозофосфатов
Реакции ПЕНТОЗОФОСФАТНОГО ЦИКЛА И ФЕРМЕНТЫИХ КАТАЛИЗИРУЮЩИЕ
| Номер р-ции на схеме | реакции | Фермент, катализирующий р-цию | ||
6 Глюкозо-6-фосфат + 6 НАДФ 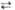 6 6-Фосфоглюко-нолактон + 6 НАДФН + 6Н+ 6 6-Фосфоглюко-нолактон + 6 НАДФН + 6Н+
| Глюкозо-6-фос- фат-дегидроге-наза | |||
6 6-Фосфоглюконолактон  6 6-Фосфоглюконат 6 6-Фосфоглюконат
| Лактоназа | |||
6 6-Фосфоглюконат + 6 НАДФ 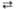 6 Риболозо-5-фосфат + 6 НАДФН + 6 H+ + 6CO2 6 Риболозо-5-фосфат + 6 НАДФН + 6 H+ + 6CO2
| 6-Фосфоглюко-нат-дегидроге-наза | |||
2 Рибулозо-5-фосфат 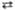 2 Рибозо-5-фосфат 2 Рибозо-5-фосфат
| Пентозоизоме-раза | |||
4 Рибулозо-5-фосфат 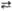 4 2-Ксилулозо-5-фосфат 4 2-Ксилулозо-5-фосфат
| Фосфорибулозо-эпимераза | |||
2 Ксилолуозо-5-фосфат + 2 Рибозо-5-фосфат 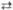 2 Седогептулозо-7-фосфат + 2 Глиперальдегид-3-фосфат 2 Седогептулозо-7-фосфат + 2 Глиперальдегид-3-фосфат
| Транскетолаза | |||
2 Седогептулозо-7-фосфат + 2 Глицеральдегид-3-фосфат 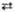 2 Эритрозо-4-фосфат + 2 Фруктозо-6-фосфат 2 Эритрозо-4-фосфат + 2 Фруктозо-6-фосфат
| Трансальдолаза | |||
2 Ксилулозо-5-фосфат + 2 Эритрозо-4-фосфат 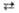 2 Глицеральдегид-3-фосфат + 2 Фруктозо-6-фосфат 2 Глицеральдегид-3-фосфат + 2 Фруктозо-6-фосфат
| Транскетолаза | |||
Глицеральдегид-3-фосфат 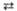 Дигидроксиацетон-фосфат Дигидроксиацетон-фосфат
| Триозофосфат-изомераза | |||
Дигидроксиацетонфосфат + Глицеральдегид-3-фос-фат 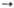 Фруктозо- 1,6-дифосфат Фруктозо- 1,6-дифосфат
| Альдолаза | |||
Фруктозе- 1, 6- дифосфат 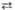 Фруктозо-6-фосфат +
+ Фруктозо-6-фосфат +
+ 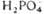
| Фруктозо-бис-фосфатаза | |||
5 Фруктозо-6-фосфат 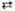 5 Глюкозо-6-фосфат 5 Глюкозо-6-фосфат
| Гексозофосфат-изомераза | |||
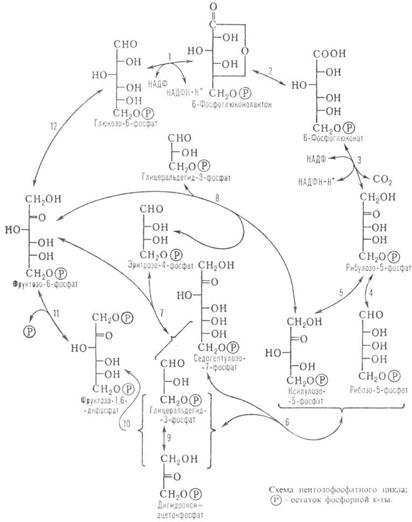
ПЕНТОЗОФОСФАТНЫЙ ЦИКЛ (пентозный путь, гексо-зомонофосфатный шунт, фосфоглюконатный путь), совокупность обратимых ферментативных р-ций, в результате к-рых происходит окисление глюкозы до CO2 с образованием восстановленного никотинамидадениндинуклеотид-фосфата (НАДФН) и H +, а также синтез фосфорилир. Сахаров, содержащих от 3 до 7 атомов С.
Пентозофосфатный цикл осуществляется в цитозоле (жидкой фазе) клеток животных, растений (особенно в темноте) и микроорганизмов. У растений часть р-ций пентозофосфатного цикла участвует также в образовании гексоз при фотосинтезе.
15)глюконеогенез — процесс образования в печени и отчасти в корковом веществе почек (около 10 %) молекул глюкозы из молекул других органических соединений — источников энергии, например свободныхаминокислот, молочной кислоты, глицерина. Свободные жирные кислоты у млекопитающих для глюконеогенеза не используются.
1. Стадии глюконеогенеза повторяют стадии гликолиза в обратном направлении и катализируются теми же ферментами за исключением 4 реакций: Превращение пирувата в оксалоацетат (фермент пируваткарбоксилаза)
2. Превращение оксалоацетата в фосфоенолпируват (фермент фосфоенолпируваткарбоксикиназа)
3. Превращение фруктозо-1,6-дифосфата в фруктозо-6-фосфат (фермент фруктозо-1,6-дифосфатаза)
4. Превращение глюкозо-6-фосфата в глюкозу (фермент глюкозо-6-фосфатаза)
Суммарное уравнение глюконеогенеза: 2 CH3COCOOH + 4ATP + 2GTP + 2NADH.H+ + 6 H2O = C6H12O6 + 2NAD + 4ADP + 2GDP + 6Pn
При голодании в организме человека активно используются запасы питательных веществ (гликоген, жирные кислоты). Они расщепляются доаминокислот, кетокислот и других неуглеводных соединений. Большая часть этих соединений не выводится из организма, а подвергаются реутилизации. Вещества транспортируются кровью в печень из других тканей, и используются в глюконеогенезе для синтеза глюкозы — основного источника энергии в организме. Таким образом при истощении запасов организма, глюконеогенез является основным поставщиком энергетических субстратов.
16) Са́харный диабе́т (лат. diabetes mellītus) — группа эндокринных заболеваний, развивающихся вследствие абсолютной или относительной (нарушение взаимодействия с клетками-мишенями) недостаточности гормона инсулина, в результате чего развивается гипергликемия — стойкое увеличение содержания глюкозы в крови. Заболевание характеризуется хроническим течением и нарушением всех видов обмена веществ: углеводного, жирового, белкового, минерального и водно-солевого