ПАЛЕОНТОЛОГИЧЕСКИЕ ДОКАЗАТЕЛЬСТВА ЭВОЛЮЦИИ
Палеонтология — наука, изучающая ископаемые остатки организмов. Палеонтологические доказательства эволюции:
1) Ископаемые остатки и отпечатки (окаменелости) древних организмов показывают, как шло их историческое развитие (эволюция). Ископаемые остатки организмов (окаменелости, фоссилии) - остатки или следы жизнедеятельности организмов геол. прошлого, сохранившиеся в осадочных породах. Лучше сохраняются в водных бассейнах: на дне платформенных морей, озёр, болот и торфяников, в отложениях рек, особенно в их дельтах, где организмы после гибели покрываются осадком, который предохраняет их от полного разрушения. Чем тонкозернистее осадок, тем детальнее отпечатывается на нём поверхность организма (напр., тончайшие жилки листьев кайнозойских растений или особенности скульптуры раковин моллюсков). Именно поэтому в породах мор. происхождения часто встречаются окаменелости. В наземных условиях большинство животных и растений обычно полностью разрушается. Различают несколько форм сохранности ископаемых остатков. Полная сохранность организмов, в т. ч. и мягких тканей, возможна в редких случаях, например, при захоронении в вечной мерзлоте (трупы мамонтов, лошадей), в озокерите — минер, воскоподобном веществе (птицы, насекомые), при естественной мумификации. Особая форма сохранности — инклюзы в меловых и палеогеновых смолах — янтаре и сходных с ним образованиях (насекомые и др. членистоногие, части растений). Растения, особенно крупные, как правило, не сохраняются целиком; от них остаются разрозненные листья, обрывки ветвей, обломки стволов, шишки, плоды, отд. семена, споры, пыльца, редко — цветки. От животных сохраняются преимущественно части минерализованного скелета, в т. ч. раковины, панцири и зубы. Ископаемые остатки могут сохранять свой первичный состав или минерализоваться в процессе фоссилизации (окаменения), т. е. замещения твёрдых (реже мягких) частей организма минер, веществами. Замещающими минералами могут быть пирит, кварц, углекислый кальций и др. Окаменение может происходить также путём преобразования первичной структуры скелета без изменения его химического состава (например, превращение арагонита в кальцит) и заполнения пустот в скелетных остатках различными минеральными веществами, растворёнными в воде. Часто от организма остаётся лишь отпечаток — запечатлённая в осадке форма твёрдых и мягких частей тела животного или растения. Распространённая форма ископаемых остатков — ядро — состоящий из породы слепок полости, образовавшийся в породе на месте организма при его разрушении. При заполнении внутренней полости скелета или раковины осадком и при последующем разрушении этого скелета образуется внутреннее ядро. При разрушении раковины или скелета и заполнении оставшейся полости минер, веществом образуется внеш. ядро, передающее особенности наружного строения организма. Своеобразная форма сохранности — следы жизни. Нередко ископаемые остатки полностью слагают породу (ракушечники, нуммулитовые, коралловые, водорослевые известняки и др. отложения). Некоторые ископаемые остатки используются для установления относительного возраста вмещающих их пород.

| 
| 
| 
| 
|
| Фоссилии тарбозавра | Морские фоссилии | Аммониты. Окаменелые раковины | Трилобит | Отпечаток папоротника |
2) Филогенетические ряды – это ряды видов, последовательно сменявших друг друга в процессе эволюции.
Ученые-палеонтологи, опираясь на идеи Дарвина, исследовали историю развития животных. Владимир Онуфриевич Ковалевский (1842-1883) в основном занимался исследованием парнокопытных животных. Он, изучив причины эволюционного процесса на примере лошади, сделал вывод о том, что изменения в ее строении связаны с изменением среды обитания. С изменением климата уменьшались лесные массивы, а увеличение открытых степных пространств способствовало изменению жизненной среды предков лошадей. Они стали приспосабливаться к условиям степей. Для спасения от врагов и поиска пастбищ с богатой растительностью необходимо было научиться быстро бегать. Перемещения на большие расстояния способствовали изменению числа пальцев на конечностях. Изменениям подверглись не только конечности, но и форма тела, черепа, строение зубов — в связи с пережевыванием травы. Произошла полная перестройка организма лошади. Такими изменениями явились: уменьшение количества пальцев от пяти до одного, укрепление тела, усложнение коренных зубов. Все это свидетельствует о процессе эволюции лошади. Итак, постепенные переходы одних видов в другие, одних родов в другие — доказательство эволюционного процесса.
В. О. Ковалевский изучал эволюцию современных лошадей и назвал ступени их развития названиями веков третичного периода кайнозойской эры. Лошадей расположил по мере уменьшения числа пальцев на конечности.
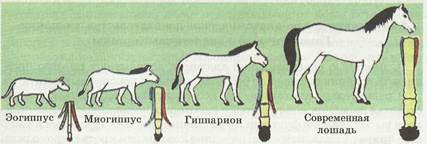
| 1. Фенакодус (существовал в палеоцене), пятипалый, ростом с лисицу. 2. Эогиппус (жил в эоцене), рост 30 см, с четырехпалыми передними конечностями, трехпалыми задними. 3. Миогиппус (жил в миоцене), хорошо развит средний палец, второй и четвертый пальцы короткие. 4. Парагиппус (жил в миоцене), сильно развит средний палец, второй и четвертый пальцы укорочены. 5. Плиогиппус (существовал в плиоцене), однопалый, остальные пальцы атрофированы. 6. Современная лошадь. |
Открытие В. О. Ковалевским систематического ряда в изучении видов ископаемых лошадей — типичный пример эволюционного процесса. Наличие последовательно сменяющих друг друга форм позволяет построить ряд с высокой степенью достоверности эволюции, который называется филогенетическим.
В результате изучения ископаемых остатков парнокопытных млекопитающих В. О. Ковалевский и его преемники выяснили историю развития лошади (филогенез).
Гильгендорф (1866) описал палеонтологический ряд брюхоногих моллюсков из миоценовых отложений, накопившихся в течение двух миллионов лет в озерных отложениях Штейнгеймского бассейна (Вюртемберг, Германия). Было обнаружено в последовательных слоях 29 различных форм, принадлежащих к ряду планорбис (Planorbis). Древние моллюски имели раковину в виде спирали, а более поздние — в виде турбоспирали. Ряд имел два ответвления. Предполагается, что изменение формы раковины было вызвано повышением температуры и увеличением содержания карбоната кальция в горячих вулканических источниках.
Таким образом, филогенетические ряды представляют собой историческую последовательность переходных форм.
В настоящее время известны филогенетические ряды для аммонитов (Вааген, 1869), брюхоногих моллюсков рода живородок (Viviparus) (Неймайром, 1875), носорогов, слонов, верблюдов, парнокопытных и других животных.
3) Переходные формы — организмы, которые сочетают в своем строении признаки двух больших систематических групп. Переходные формы характеризуются наличием более древних и примитивных (в смысле первичных) черт, чем более поздние формы, но, в то же время, наличием более прогрессивных (в смысле более поздних) черт, чем их предки. Как правило, термин «переходные формы» употребляют по отношению к ископаемым формам, хотя промежуточные виды совсем не обязательно должны умирать. Переходные формы используют как одно из доказательств существования биологической эволюции.
В 1859 г., когда была издана работа Ч. Дарвина «Происхождение видов», количество ископаемых остатков была крайне мало, науке не были известны переходные формы. Дарвин описал отсутствие промежуточных форм «как наиболее очевидное и тяжелое возражение, которое может быть против теории», но объяснил это крайней неполнотой геологической летописи. Он отмечал ограниченное количество доступных коллекций, в то же время, описал имеющуюся информацию об имеющихся ископаемые образцы с точки зрения эволюции и действия естественного отбора. Только два года спустя, в 1961 году, был найден археоптерикс, который представлял классическую переходную форму между пресмыкающимися и птицами. Его находка стала не только подтверждением теории Дарвина, а также знаковым фактом, подтверждающий реальность существования биологической эволюции. С тех пор было найдено большое количество ископаемых форм, которые показывают, что все классы позвоночных животных являются родственными между собой, причем большинство из них — через переходные формы. С увеличением сведений о таксономическом разнообразии сосудистых растений в начале ХХ в., начались исследования по поиску их возможного предка. В 1917 г. Роберт Кидстон и Уильям Генри Ленд обнаружили остатки очень примитивной растения возле поселка Rhynia в Шотландии. Это растение было названо Rhynia (риниофит). Она сочетает в себе признаки зеленых водорослей и сосудистых растений.
Переходные формы между двумя группами организмов не обязательно потомками одной группы и предком другой. По ископаемым, как правило, невозможно точно установить является ли определенный организм предком другого. Кроме того, вероятность найти в палеонтологической летописи прямого предка определенной формы чрезвычайно мала. Гораздо больше вероятность обнаружить относительно близких родственников этого предка, которые сходны с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок филогенетического ствола».
Отсутствие переходных форм между многими группами организмов является предметом критики со стороны креационистов. Однако далеко не каждая переходная форма существует в виде окаменелостей из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фоссилизации, то есть перехода в окаменевшее состояние. Для образования окаменелости необходимо, чтобы организм, который погиб, был погребен под большим слоем осадочных пород. Из-за очень медленной скорости осадконакопления на суше, сухопутные виды редко переходят в окаменевшее состояние и сохраняются. Кроме того, редко удается выявить виды, которые живут в глубинах океана через редкие случаи поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, или наземные виды, которые ведут полуводный образ жизни, или живут у береговой линии. К упомянутым выше проблемам следует добавить чрезвычайно малое (в масштабах планеты) количество палеонтологов, которые осуществляют раскопки.
Переходные формы, как правило, не живут на больших территориях и не существуют в течение большого времени. Этот факт также снижает вероятность фоссилизации и последующего обнаружения переходных форм. Поэтому вероятность обнаружения промежуточных форм чрезвычайно мала.

| 
|  Археоптерикс
Археоптерикс
|  Конфуциусорнис
Конфуциусорнис
| |||
| Древнейшими представителями земноводных считают ихтиостеги. Их считают переходным звеном между кистеперых рыбами и земноводными. Несмотря на то, что в ихтиостеги была пятипалая концовка, адаптированная к жизни на суше, значительную часть жизни они проводили как рыбы, имели хвостовой плавник, боковую линию и некоторые другие признаки рыб. | Батрахозавры, существовавшие в каменноугольный и пермский периоды, рассматривают как переходную форму между земноводными и пресмыкающимися. Батрахозавры, хоть и проводили жизнь во взрослой стадии на суше (подобно пресмыкающихся), были тесно связаны с водоемами и сохранили ряд признаков, присущих земноводным, в частности, откладывания икры и развитие личинок в воде, наличие жабр и тому подобное. | Обнаружено большое количество пресмыкающихся, которые выработали способность летать, часть из них имела перья, поэтому их рассматривают как переходные формы между пресмыкающимися и птицами. Наиболее известен археоптерикс. Он был размером примерно с современную ворону. Формой тела, строением конечностей и наличием оперения подобен современным птицам, возможно, летал. Общим с пресмыкающимися была особое строение таза и ребер, наличие клюва с коническими зубами, по три свободных пальца на крыльях, длинный хвост из 20-21 позвонка, кости не пневматизованные, грудина без киля. Другие известные переходные формы между пресмыкающимися и птицами — протоавис, конфуциусорниса. | ||||

| 
|  Лиценопс
Лиценопс
|  Иностранцевия
Иностранцевия
| |||
| Амбулоцетус (Ambulocetus natans, «ходячий кит») — переходная форма между наземными млекопитающими и китообразными, которые являются вторичноводными. Внешне животное напоминало нечто среднее между крокодилом и дельфином. Кожа с частично редуцированной шерстью. Животное имело лапы с перепонками; хвост и конечности приспособлены как вспомогательные органы передвижения в воде. | Первые наземные растения из класса риниопсид, семей риниевих и псилофитовых, живших в силуре — девоне, сочетали признаки зеленых водорослей и примитивных форм высших растений. Их тело было безлистным, цилиндрический осевой орган — телом в верхней части дихотомически разветвленным на верхушках с спорангиями. Функцию минерального питания риниопсид выполняли ризоиды. | Большое количество ископаемых форм звероподобных пресмыкающихся (синапсиды, терапсид, пеликозавров, различных динозавровых и др.), найденных во многих районах земного шара, существовали в юрский и меловой периоды, сочетающие признаки пресмыкающихся и млекопитающих, раскрывают возможные направления и способы становления различных групп четвероногих, в частности млекопитающих. Например, звероподобное пресмыкающееся из группы терапсид — лиценопс (Lycaenops) по развитию костей ротовой полости, дифференцированием зубов на клыки, резцы, по резцовые зубы и рядом других признаков строения тела напоминает хищных млекопитающих, хотя по другим признакам и образом жизни это были настоящие пресмыкающиеся. | ||||

| Ископаемые формы семенных папоротников, которые процветали в конце девона, сочетают в себе признаки папоротников и голосеменных. Они образовывали не только споры (как папоротники), но и семена (как семенах растения). Проводящая ткань их стеблей по строению напоминает древесину голосеменных (саговников). Рисунок Семенные папоротники: 1 - реконструкция семенного папоротника из группы медуллозовых (род Medullosa); 2 - пёрышки папоротника Pecopteris с семенами; 3 - лист Neuropteris с семенем; 4 - реконструкция семени Lagenostoma Iomaxii, принадлежащего лигиноптерис; А - семена, К - купула. | 
| Другой предшественник семенных растений был идентифицирован из отложений среднего девона. Рункария (Runcaria heinzelinii) существовала около 20 млн лет назад. Найденные ископаемые остатки Runcaria heinzelinii представляют собой короткие разветвленные стебли, на концах которых имеется радиально-симметричный мегаспорангий, окруженный купулой. Мегаспорангий имел многолепестковые покровы и расширение, которое предлагает адаптацию к опылению ветра. Таким образом, это ископаемое растение обладает всеми качествами семенных растений, за исключением твердого семенного покрова и системы для направления пыльцы к яйцеклетке. Это проливает новый свет на то, как семя могло эволюционировать. | |||