Лекция 3.
ЭКОСИСТЕМЫИ ПРИНЦИПЫИХ ФУНКЦИОНИРОВАНИЯ
Живые организмы в биоценозах тесно связаны не только друг с другом, но и с неживой природой через вещество и энергию. Протекающие через живые организмы потоки вещества и энергии в процессе обмена веществ весьма велики. Человек, например, за свою жизнь потребляет десятки тонн пищи и воды, тысячи кубометров воздуха.
Чрезвычайно высокая интенсивность потоков вещества из неорганической природы в живые тела давно привела бы к полному исчерпанию запасов необходимых для жизни соединений, то есть биогенных элементов. Но этого не происходит, и жизнь не прекращается, так как указанные элементы постоянно возвращаются в окружающую среду. И происходит это благодаря биоценозам, в которых в результате пищевых отношений между видами синтезированные растениями сложные органические вещества превращаются в конце концов в такие простые соединения, как диоксид углерода, вода, ряд элементов, которые могут быть снова использованы растениями в процессе фотосинтеза. Так возникает биологический круговорот вещества. Следовательно, биоценоз, будучи и сам по себе сложной системой живых организмов, является частью еще более сложной системы. В последнюю, помимо живых организмов, входит и их неживое окружение, которое содержит различные вещества и энергию, необходимые для развития и обеспечения жизнедеятельности.
Любую совокупность организмов и неорганических компонентов окружающей их среды, в которой может осуществляться круговорот веществ, называют экологической системой, или экосистемой.
Или
Экологическая система (экосистема) - совокупность популяций различных видов растений, животных и микробов, взаимодействующих между собой и окружающей их средой таким образом, что эта совокупность сохраняется неопределенно долгое время.
Природные экосистемы могут быть самого различного объема и протяженности. Это и капля воды с ее обитателями, и лужа, пруд, луг, тайга, степь (рис. 2.1).

Рис. 2.1. Группы природных экосистем
Однако любая экосистема, независимо от размера, включает в себя живую часть (биоценоз) и ее физическое, то есть неживое, окружение. При этом малые экосистемы входят в состав все более крупных, вплоть до глобальной экосистемы Земля. Аналогично общий биологический круговорот вещества на планете также складывается из взаимодействия множества более мелких, частных круговоротов.
Экосистема может обеспечить круговорот веществ только в том случае, если включает четыре необходимые для этого части:
1) запасы биогенных элементов;
2) продуценты;
3) консументы;
4) редуценты.
На их сложном и постоянном взаимодействии основан первый (основной) принцип функционирования экосистем: получение ресурсов и избавление от отходов происходят в рамках круговорота всех элементов.
Данный принцип гармонирует с законом сохранения массы. Так как атомы не возникают, не исчезают и не превращаются один в другой, они могут использоваться бесконечно в самых различных химических соединениях, и запас их практически неограничен. Именно это и происходит в природных экосистемах.
Необходимо подчеркнуть, однако, что биологический круговорот не совершается исключительно за счет вещества, поскольку он — результат деятельности организмов, для обеспечения жизнедеятельности которых требуются постоянные энергетические затраты, поставляемые Солнцем. Энергия солнечных лучей, поглощаемая зелеными растениями, в отличие от химических элементов, не может использоваться организмами бесконечно. Данное заключение вытекает из второго закона термодинамики: энергия при превращении из одной формы в другую, то есть при совершении работы, частично переходит в тепловую форму и рассеивается в окружающей среде.
Следовательно, каждый цикл круговорота, зависящий от активности организмов и сопровождаемый потерями энергии из них, требует все новых дотаций энергии. Существование экосистем любого ранга и вообще жизни на Земле обусловлено постоянным круговоротом веществ, который, в свою очередь, поддерживается постоянным притоком солнечной энергии. В этом состоит второй основной принцип функционирования экосистем: они существуют за счет не загрязняющей среду и практически вечной солнечной энергии, количество которой относительно постоянно и избыточно.
ПОТОКИ ВЕЩЕСТВА И ЭНЕРГИИ В ЭКОСИСТЕМЕ
Предварительно необходимо определиться в этих терминах.
Поток вещества — перемещение последнего в форме химических элементов и их соединений от продуцентов к редуцентам (через консументы или без них).
Поток энергии — переход энергии в виде химических связей органических соединений (пищи) по цепям питания от одного трофического уровня к другому (более высокому).
Подчеркнем тот факт, что, в отличие от веществ, которые постоянно циркулируют по разным блокам экосистемы и всегда могут вновь входить в круговорот, поступившая энергия может быть использована только один раз.
Как универсальное явление природы, односторонний приток энергии обусловлен действием законов термодинамики. Согласно первому из них: энергия может переходить из одной формы (энергии света) в другую (потенциальную энергию пищи), но она никогда не создается вновь и не исчезает бесследно.
Согласно второму закону термодинамики, не может быть ни одного процесса, связанного с превращением энергии, без потери некоторой ее части. Поэтому не может быть превращений, например, пиши в вещество, из которого состоит тело организма, идущих со 100%-ной эффективностью.
Таким образом, функционирование всех экосистем определяется постоянным притоком энергии, которая необходима всем организмам для поддержания их существования и самовоспроизведения.
В процессе изучения развития экосистем следует учитывать и конкурентные отношения. В этом аспекте большой интерес представляет закон максимизации энергии: в соперничестве с другими экосистемами выживает (сохраняется) та из них, которая наилучшим образом способствует поступлению энергии и использует максимальное ее количество наиболее эффективным способом.
С этой целью система:
1) создает накопители (хранилища) высококачественной энергии (например, запасы жира);
2) затрачивает определенное количество накопленной энергии на обеспечение поступления новой энергии;
3) обеспечивает круговорот различных веществ;
4) создает механизмы регулирования, поддерживающие устойчивость системы и ее способность к приспособлению к изменяющимся условиям;
5) налаживает с другими системами обмен, необходимый для обеспечения потребности в энергии специальных видов».
Закон максимизации энергии справедлив и в отношении информации, следовательно, его возможно рассматривать и как закон максимизации энергии и информации с такой формулировкой: наилучшими шансами на самосохранение обладает система, в наибольшей степени способствующая поступлению, выработке и эффективному использованию энергии и информации.
Ранее отмечалось, что между организмами биоценоза возникают и устанавливаются прочные пищевые взаимоотношения или цепь питания. Последняя состоит из трех основных звеньев: продуцентов, консументов и редуцентов.
Цепи питания, которые начинаются с фотосинтезирующих организмов, называют цепями выедания (или пастбищными), а цепи, начинающиеся с отмерших остатков растений, трупов и экскрементов животных, — детритными цепями.
Место каждого звена в цепи питания называют трофическим уровнем, он характеризуется различной интенсивностью протекания потока веществ и энергии.
Первый трофический уровень всегда составляют продуценты; растительноядные консументы относятся ко второму трофическому уровню; плотоядные, живущие за счет растительноядных форм - к третьему; потребляющие других плотоядных — соответственно к четвертому и т.д. Вследствие этого различают консументов первого, второго, третьего и четвертого порядков, занимающих разные уровни в цепях питания. Упрощенная схема цепи питания представлена на рис. 2.2.
Очевидно, что основную роль при этом играет пищевая специализация консументов. Виды с широким спектром питания могут включаться в цепи питания на разных трофических уровнях. В рацион, например, человека входит как растительная пища, так и мясо травоядных и плотоядных животных. Поэтому он выступает в разных пищевых цепях в качестве консумента первого, второго или третьего порядков.
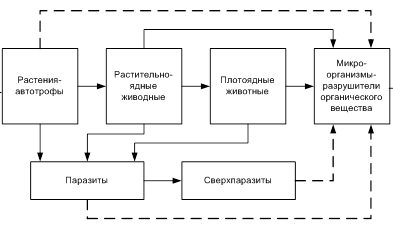
Рис 2.2. Упрощенная схема питания
Так как при передаче энергии с одного уровня на другой происходит ее потеря, цепь питания не может быть длинной. Обычно она состоит из 4—6 звеньев.
Однако такие цепи в чистом виде в природе обычно не встречаются, поскольку одни и те же виды могут быть одновременно в разных звеньях. Это обусловлено тем, что монофагов в природе мало, намного чаще встречаются олигофаги и полифаги. Например, хищники, которые питаются различными растительноядными и плотоядными животными, являются звеньями многих цепей. Из-за этого в каждом биоценозе исторически формируются комплексы цепей питания, представляющие собой единое целое. Подобным образом создаются сети питания, которые отличаются большой сложностью.
Таким образом, можно сделать вывод о том, что пищевая цепь — основной канал переноса энергии в сообществе.
На рис. 2.3 показана схема переноса энергии между растениями-продуцентами, животными-консументами и микроорганизмами-редуцентами.
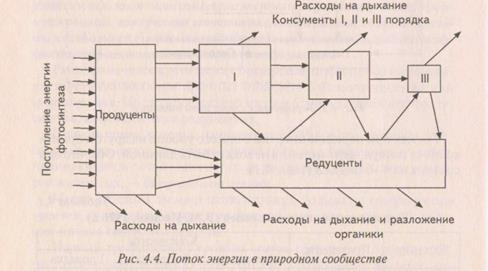
Рис. 2.3. Поток энергии в природном сообществе
Уже из этой схемы видно, что представление о пищевых цепях и трофических уровнях — скорее абстракция. Линейную цепь с четко разделенными уровнями можно создать в лабораторных условиях. Однако в природе реально существуют трофические сети, в которых многие популяции принадлежат сразу к нескольким трофическим уровням. Один и тот же организм потребляет в пищу и животных, и растения; хищник может питаться консументами /и //порядка; многие животные едят как живые, так и отмершие растения.
Благодаря сложности трофических связей выпадение какого-то одного вида нередко почти не сказывается на сообществе. Пищу исчезнувшего вида начинают потреблять другие «пользователи», питавшиеся им виды находят новые источники пищи, и в целом в сообществе сохраняется равновесие.
Теперь рассмотрим, как и в каком соотношении передается энергия, заключенная в растительной пище, по цепям питания.
В ходе фотосинтеза растения связывают в среднем лишь около 1 % попадающей на них солнечной энергии. Животное, которое съело растение, часть пищи не переваривает и выделяет в виде экскрементов. Усваивается обычно 20-60% растительного корма, усвоенная энергия расходуется на поддержание жизнедеятельности животного. Функционирование организма сопровождается выделением тепла, в результате существенная доля энергии пищи вскоре рассеивается в окружающей среде. Сравнительно небольшая часть пищи идет на построение новых тканей и создание жировых запасов. В дальнейшем хищник, съевший это растительноядное животное и представляющий третий трофический уровень, получает только ту энергию из накопленной растением, которая задержалась в теле его жертвы (второй уровень) в виде прироста биомассы.
Согласно расчетам, на каждом этапе передачи вещества и энергии по пищевой цепи теряется примерно 90% энергии и только около одной десятой доли ее переходит к очередному потребителю. Указанное соотношение в передаче энергии в пищевых связях организмов называют «правилом десяти процентов»
Например, количество энергии, которая доходит до третичных плотоядных (пятый трофический уровень), составляет лишь около 10~4 энергии, поглощенной продуцентами. Тем самым объясняется ограниченное количество (5—6) звеньев (уровней) в пищевой цепи независимо от сложности видового состава биоценоза.
Рассматривая поток энергии в экосистемах, легко понять также, почему с повышением трофического уровня биомасса снижается. Здесь проявляется третий основной принцип функционирования экосистем: чем больше биомасса популяции, тем ниже должен быть занимаемый ею трофический уровень, или иначе: на конце длинных пищевых цепей не может быть большой биомассы.
ДИНАМИКА ЭКОСИСТЕМ
Биоценозы, независимо от их сложности, динамичны, в них постоянно происходят изменения в состоянии и жизнедеятельности их членов и соотношении популяций. Указанные изменения можно свести к двум основным типам: циклическим и поступательным.
Циклический тип изменения сообществ отражает суточную, сезонную и многолетнюю периодичность внешних условий и проявления эндогенных (внутренних) ритмов организмов.
В любом естественном биоценозе имеются группы организмов, активность жизни которых приходится на разное время суток. Поэтому в составе и соотношении отдельных видов биоценоза происходят периодические изменения, так как ряд организмов на определенное время выключаются из него. При этом суточную динамику биоценоза обеспечивает не только животное, но и растительное население.
Суточная динамика биоценозов преимущественно связана с ритмами природных явлений и характеризуется строгой периодичностью.
При сезонной динамике имеют место более существенные отклонения в биоценозах, определяемые биологическими циклами организмов, которые зависят от сезонной цикличности природных явлений. Смена времен года существенно влияет на жизнедеятельность растений и животных (периоды цветения, плодоношения, активного роста, осеннего листопада и зимнего покоя у растений; спячка, зимний сон, диапауза и миграции у животных).
Нормальным явлением в жизни любого биоценоза служит и многолетняя изменчивость. Она обусловлена изменением по годам метеорологических условий (климатических флюктуации) или других внешних факторов, которые влияют на сообщество (например, разливы рек). Кроме этого, многолетняя периодичность часто бывает связана с особенностями жизненного цикла растений - эдификаторов, с повторением массовых размножений животных, насекомых или патогенных для растений микроорганизмов.
Поскольку характер суточных и сезонных изменений более или менее постоянен в течение длительного периода времени (столетий и даже тысячелетий), исторически сформировались механизмы, которые приводят сообщество в целом в соответствие с периодикой изменения условий обитания.
Поступательные изменения в сообществе приводят в конечном итоге к смене этого сообщества другим, с иным набором господствующих видов. Причиной подобных смен могут быть внешние по отношению к биоценозу факторы, длительное время воздействующие в одном направлении, например, иссушение болотных почв.
БИОЛОГИЧЕСКАЯ ПРОДУКТИВНОСТЬ ЭКОСИСТЕМ. ПЕРВИЧНАЯ И ВТОРИЧНАЯ ПРОДУКЦИЯ.
Скорость создания органического вещества в экосистемах называется биологической продукцией, а масса тела живых организмов — биомассой. Следовательно, биологическая продукция экосистем — это скорость создания в них биомассы.
Органическая масса, создаемая растениями за единицу времени, называется первичной продукцией сообщества, а продукция животных или других консументов — вторичной. Очевидно, что вторичная продукция не может быть больше первичной или даже равной ей. Продукцию выражают количественно в сырой или сухой массе растений либо в энергетических единицах — эквивалентном числе джоулей.
Валовая первичная продукция — количество вещества, которое создается растениями за единицу времени при данной скорости фотосинтеза. Первичную биологическую продукцию экосистем ограничивают или неблагоприятные климатические факторы (недостаток тепла, влаги), или нехватка биогенных элементов. Можно привести примеры продуктивности различных экосистем (в граммах сухого вещества на квадратный метр площади за сутки): менее 1 г — пустыни, глубокие моря; 1—3 г — луга, горные леса, пашни, мелкие моря, глубокие озера; 3—10 г — степи, мелкие озера, леса умеренной полосы, орошаемые поля; 10—25 г — тропические леса, интенсивно возделываемые культуры на полях, коралловые рифы.
Часть производимой продукции идет на поддержание жизнедеятельности самих растений (затраты на дыхание). В тропических лесах и зрелых лесах умеренной полосы она составляет 40—70% валовой продукции. Около 40% составляют затраты на дыхание у большинства сельскохозяйственных культур.
Оставшаяся часть созданной органической массы характеризует чистую первичную продукцию. Представляя собой величину прироста биомассы растений, она является энергетическим резервом для консументов и редуцентов. Постепенно перерабатываясь в цепях питания, она идет на пополнение биомассы гетеротрофных организмов.
Правило пирамид. Всем экосистемам отвечают определенные соотношения первичной и вторичной продукции, называемые правилом пирамиды продукции: на каждом предыдущем трофическом уровне количество биомассы, создаваемой за единицу времени, больше, чем на последующем.
Например, масса всех трав, выросших за год в степи, значительно больше, чем годовой прирост всех растительноядных животных, а прирост хищников меньше, чем растительноядных животных.
Указанное правило отображают в виде пирамид, сужающихся кверху и образованных поставленными друг на друга прямоугольниками равной высоты. Длина этих прямоугольников соответствует масштабам продукции на соответствующих трофических уровнях (рис. 4.5).
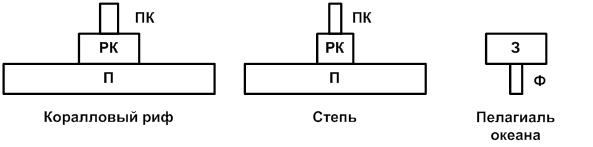
Рис. 5 Пирамиды биомассы некоторых сообществ, где:
П – продуценты; РК – растительноядные консументы; ПК – плотоядные консументы; Ф – фитопланктон; З – зоопланктон (крайняя справа пирамида биомассы имеет перевернутый вид)
Известно, что основными продуцентами в океане являются одноклеточные водоросли, отличающиеся высокой скоростью оборота генераций. Как следствие, их годовая продукция может в десятки и даже сотни раз превышать запас биомассы на данный момент времени. Вся чистая первичная продукция так быстро вовлекается в цепи питания, то есть поедается, что накопление биомассы водорослей весьма мало. Тем не менее из-за высоких темпов размножения небольшой их запас вполне достаточен для поддержания скорости воссоздания органического вещества. Поэтому для океана пирамида биомасс имеет перевернутый вид. На высших трофических уровнях преобладает тенденция к накоплению биомассы, поскольку длительность жизни крупных хищников (например, кита-касатки) велика, скорость оборота этих генераций (поколений), наоборот, мала, и в их телах задерживается значительная часть вещества, поступающего по цепям питания.
В тех трофических цепях, где передача происходит в основном через связи «хищник-жертва», справедливо правило пирамиды чисел: общее число особей, которые участвуют в цепях питания, с каждым последующим звеном уменьшается.
Поясним это правило. Хищник обычно крупнее своих жертв, и для поддержания собственной биомассы ему нужно несколько или много жертв. Однако бывают случаи, когда более мелкие хищники живут за счет групповой охоты на крупных животных.
Подчеркнем, что из правила пирамиды биологической продукции нет исключений, потому что оно отражает законы передачи энергии в цепях питания.
Пирамида энергии более точно отображает трофические связи организмов, поскольку она характеризует скорость возобновления биомасс. На каждом уровне пирамида энергии отражает удельное количество энергии (на единицу площади или объема), прошедшей через предыдущий трофический уровень за данный отрезок времени. Пирамиды потоков энергии никогда не бывают «перевернутыми»: следующий трофический уровень может «пропустить через себя» лишь часть энергии, усвоенной предыдущим уровнем (вспомните правило 10%).
Изучение законов продуктивности экосистем, возможность количественного учета потока энергии чрезвычайно важны в практическом отношении, так как первичная продукция агроценозов и эксплуатируемых человеком природных сообществ — основной источник запасов пищи для человечества. Весьма важна и вторичная продукция, которую получают за счет сельскохозяйственных и промышленных животных: животные белки содержат целый ряд незаменимых для человека аминокислот, которых нет в растительной пище. Точные расчеты потока энергии и масштабов продуктивности экосистем позволяют регулировать в них круговорот веществ таким образом, чтобы обеспечить наибольший выход необходимой для людей продукции. Наконец, очень важно хорошо представлять допустимые пределы изъятия растительной и животной биомассы из природных систем. В противном случае может быть подорвана их продуктивность.