ДНК (дезоксирибонуклеиновая кислота) - Дезоксирибонуклеиновая кислота представляет собой полимер, состоящий из двух полинуклеотидных цепей, которые извиваются вокруг друг друга, образуя двойную спираль, несущих генетические инструкции для развития, функционирования, роста и размножения.
Опыт Гриффитса.
Работал с двумя штаммами пневмококка, которые отличались по внешнему виду и болезнетворным свойствам. Штамм S – от него мыши сразу погибали. Штамм R – при введении гибели мышей не наступало. Он заметил, что при смеси убитых вирулетных и невирулетных мышы погибали. Это доказало процесс транформации.
Трансформация – включение чужеродной ДНК в бактериальную клетку. Это перенос наследственной информации от одной клетки прокариотов к другой посредством ДНК бактерии – донора или клетки – донора. (Гриффитс, 1928).
Опыт Херши и Чейза.
Они выяснили, что не все вирусы убивают бактериальную клетку. Существуют умеренные фаги. Они могут быть и вирулентными и объединятся с геномом бактериальной клетки. При этом попадая в другие бактерии вирусы привносят чужие генетические данные. Так доказан процесс трансдукции.
Трансдукция заключается в том, что вирусы, покидая бактериальные клетки, в которых они паразитировали, могут захватывать с собой часть их ДНК и, перемещаясь в новые клетки, передавать новым хозяевам свойства прежних.
Доказательства генетической роли ДНК были получены в ряде опытов по заражению бактериальных клеток вирусами. Бактериофаг состоит из белковой капсулы правильной геометрической формы и молекулы нуклеиновой кислоты, свернутой в виде спирали. Фаг прикрепляется своими отростками к клеточной оболочке, с помощью ферментов разрушает участок клеточной мембраны и чрез образовавшееся отверстие вводит свою ДНК в клетку и т.д.
Когда белок фага был помечен радиоактивной серой 35S, а ДНК – радиоактивным фосфором 32Р оказалось, что вновь образованные фаги содержали только радиоактивный фосфор, которым была помечена ДНК. Эти опыты наглядно показали, что генетическая информация от внедрившегося фага его потомкам передается только проникающей в клетку нуклеиновой кислотой, а не белком, содержащимся в капсуле вируса.
Опыт Френкель-Конрата.
Им удалось доказать, что чистая НК вируса табачной мозаики может заражать растения. И удалось создать гибрид из вирусов, в котором белковый футляр принадлежал одному, а НК другому вирусу. В таких случаях генетическая информация гибридов в точности повторяла генетическую информацию вируса, чья НК находилась в гибриде.
Современные представления о генетическом коде. Опыт Ниринберга. Синтез специфических белков. Репликация, транскрипция, трансляция. Транскрипция 4х-значного кода первичной генетической информации в 20-значный аминокислотный код белков. Виды и структура т-РНК, р-РНК, и-РНК. Мультимерная организация белков (гемоглобин человека:HbA, HbA2, HbS, HbF).
Генетический код – система расположения нуклеотидов в молекуле ДНК, контролирующая последовательность расположения аминокислот в молекуле белка.
Структура каждой молекулы ДНК индивидуальна и специфична, так как представляет собой кодовую форму записи биологической информации. С помощью 4 нуклеотидов в ДНК записана вся важная информация об организме, передающаяся по наследству
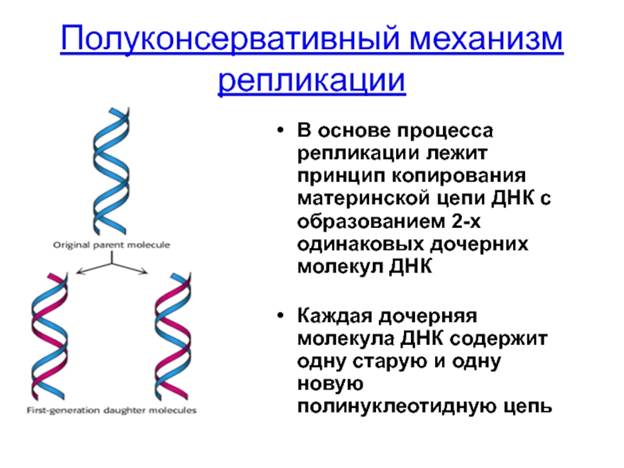
Репликация ДНК. Вспомним, что удвоение молекул ДНК, которое осуществляется в синтетическом (S) периоде клеточного цикла, называется репликацией. *После открытия Дж. Уотсоном и Ф. Криком структуры молекулы ДНК было выдвинуто несколько гипотез о возможном механизме протекания этого процесса (рис. 16.2).
Рис. 16.2.
Согласно гипотезе консервативной репликации двухцепочечная материнская молекула ДНК как единое целое служит матрицей для образования дочерней молекулы, состоящей из двух совершенно новых цепей. Полуконсервативный механизм репликации предполагает разделение цепей исходной молекулы ДНК. При этом каждая материнская цепь является матрицей для синтеза дочерней цепи. Таким образом, при репликации образуются две молекулы ДНК, каждая из которых включает одну материнскую цепь и одну новую — дочернюю. Гипотеза дисперсной репликации состояла в том, что материнская ДНК распадается на фрагменты, которые выступают в роли матриц для построения отдельных участков новых молекул. Образованные таким способом молекулы ДНК должны состоять из чередующихся фрагментов исходной молекулы и вновь синтезированных.
В 1958 г. американские биологи М. Мезельсон и Ф. Сталь провели экспериментальную проверку этих гипотез. На протяжении нескольких поколений они выращивали бактерии (а именно кишечную палочку) в питательной среде, содержащей «тяжелый» азот 15N. За это время 15N вошел в состав бактериальных молекул ДНК. Далее бактерии были перенесены в среду, содержащую изотоп 14N. Следовательно, в состав вновь синтезированных цепей ДНК включался уже «легкий» азот. Из бактериальных клеток новых поколений, образовавшихся в среде с 14N, выделяли ДНК и центрифугировали в градиенте плотности хлорида цезия (рис. 16.3).
Рис. 16.3.
Выяснилось, что ДНК дочерних клеток первого поколения имела плотность, среднюю между плотностью «легкой» ДНК, содержащей только 14N, и «тяжелой», включающей только 15N. То есть такая «гибридная» ДНК содержала одновременно 14N и 15N. Это противоречило гипотезе о консервативном механизме репликации, согласно которой ДНК должна была разделиться на две фракции — «легкую» и «тяжелую».
ДНК, выделенная из клеток второго поколения, разделялась на «легкую» и «гибридную». Этот факт полностью соответствовал гипотезе полуконсервативной репликации и позволил исключить дисперсный механизм удвоения ДНК, согласно которому плотность ДНК второго поколения бактерий должна быть средней между «легкой» и «гибридной». Таким образом был доказан полуконсервативный механизм репликации ДНК.*
*К настоящему времени установлено, что в осуществлении репликации принимает участие целый комплекс ферментов. Также известно, что этот процесс начинается в строго определенных участках молекулы ДНК — так называемых точках начала репликации. Бактериальная хромосома, как правило, имеет одну такую точку. У ядерных организмов каждая молекула ДНК (хромосома) содержит множество точек начала репликации. Следовательно, у эукариот процесс репликации одновременно протекает на многих участках одной и той же хромосомы. Это значительно сокращает время удвоения молекул ДНК.
Процесс репликации ДНК подразделяют на три этапа: инициацию, элонгацию и терминацию.
ДНК-полимераза α —первая ДНК-полимераза, обнаруженная в клетках эукариот. Она представлена в клетке в виде прочного комплекса с ДНК-праймазой — ферментом, осуществляющим синтез РНК-затравок. Комплекс ДНК-полимераза α-праймаза является единственным у эукариот ферментативным ансамблем, способным инициировать синтез ДНК de novo. В ходе репликации в клеточных ядрах ДНК полимераза α-праймаза синтезирует затравку лидирующей нити в участке ori и затравки фрагментов Оказаки запаздывающей нити. Как правило, ДНК-полимераза не обладает корректорской 3'→5'-экзонуклеазной активностью. По-видимому, в ходе эволюции экзонуклеазный центр в данном ферменте редуцировался.
ДНК-полимераза β является наименьшей по размеру и самой простой по строению ДНК-полимеразой в клетках эукариот. Основная функция ДНК-пол.β в клетке связана с эксцизионной репарацией ядерной ДНК (заполнение пробелов при репарации).
ДНК-полимераза δ —гетеродимер, состоящий из каталитической субъединицы (125—130 кДа) и субъединицы 48 — 55 кДа, необходимой для преодоления ферментом структурных барьеров в природных однонитевых матрицах и для связи с фактором процессивности PCNA (от англ. Proliferating Cell Nuclear Antigen –ядерный антиген пролиферирующих клеток). Три молекулы PCNA образуют кольцевой тример с отверстием для двунитевой ДНК в центральной части, который представляет собой перемещающуюся по ДНК подвижную платформу или «скользящую скрепку» в форме тора (бублика), удерживающую ДНК-полимеразу δ в ходе полимеризации на матрице и обеспечивающую высокопроцессивный синтез ДНК. Хотя PCNA и прокариотический фактор процессивности субъединица β ДНК-полимеразы III Е. coli имеют низкую гомологию на уровне первичной структуры, оба белка формируют близкие по пространственной геометрии структуры «скользящей скрепки».
ДНК-полимераза ε,выделена из клеток HeLa, содержит два полипептида — каталитический 261 кДа и полипептид 55 кДа. Каталитический полипептид обладает ДНК-полимеразной и 3'→5‘-экзонуклеазной активностями. Особенностью холофермента ДНК-полимеразы ε по сравнению с ДНК-полимеразой δ является его меньшая зависимость от вспомогательных факторов (PCNA, RFС - репликативный фактор С и RPA – репликативный ядерный белок А), а также низкая (почти на порядок) скорость синтеза ДНК. Это различие, возможно, связано с разной функцией ДНК-полимераз в репликативной вилке. Один холофермент, ДНК-полимераза δ осуществляет быстрый и процессивный синтез лидирующей нити, используя для элонгации единственную затравку, синтезируемую ДНК-полимеразой α-праймазой в районе ori, и диссоциирует только по достижении конца репликона, тогда как несколько холоферментов ДНК-полимеразы ε могут одновременно синтезировать фрагмены Оказаки в «зоне Оказаки», удлиняя затравки, синтезируемые ДНК-полимеразой α-праймазой в начале каждого фрагмента.
ДНК-полимераза γ локализована в митохондриях, ее функция связана с репликацией и репарацией митохондриальной ДНК, она кодируется ядерным геномом. ДНК-полимераза γ способна направлять высокопроцессивную полимеризацию на однонитевых ДНК-матрицах в отсутствие вспомогательных факторов.
Репликативная вилка
В точке репликации двойная спираль ДНК расплетается и образует петлю, к которой прикрепляются различные белки. Репликативная вилка Это расплетенная часть молекулы ДНК, где под действием специальных ферментов, происходит само удвоение цепей. Водородные связи между комплементарными азотистыми основаниями в параллельных цепях разрезаются с помощью фермента геликазы (от англ. Helix – спираль). На разрыв водородной связи расходуется энергия около двух молекул АТФ. Для того чтобы полинуклеотидные цепи не соединились вновь, к ним присоединяются специальные ДНК-связывающие белки. Распускание двойной спирали происходит очень быстро. Для того чтобы репликативная вилка могла двигаться со скоростью 500 нуклеотидов в секунду, материнская спираль должна крутиться со скоростью 50 оборотов в секунду. Так вращается вал двигателя автомобиля, который едет со скоростью 100 км/ч. Понятно, что это создает крутящий момент, который механически мешает расплетанию спирали. Многие сталкивались с таким явлением, пытаясь разделить скрученные нити. Специальный фермент гираза (от англ. Gyration – кручение), который находится перед вилкой, предотвращает запутывание расплетенных цепей. Синтез новой цепи происходит с помощью ферментного комплекса ДНК-полимеразы. Он синтезирует новую полинуклеотидную цепь, комплементарную материнской. Для понимания механизма репликации важно вспомнить о разнонаправленности и антипараллельности полинуклеотидных цепей.
Репликация у бактерий
Бактериальный геном представляет собой кольцо, размером примерно 5 млн. пар нуклеотидов. В этом кольце есть конкретное место, с которого начинается удвоение. Репликация происходит по следующему принципу. С точки начала репликации в две стороны начинают двигаться два комплекса ферментов. Каждый комплекс ферментов способен удваивать сразу обе цепочки ДНК. Область, в которой он находится, называется вилкой репликации. Два комплекса пройдут по кольцу по часовой и против часовой стрелки. Когда они встретятся, репликация будет завершена.
Ключевым ферментом, осуществляющим репликацию, является ДНК-полимераза. Она удваивает ДНК. Как правило, в каждой клетке существует несколько разных ДНК полимераз, для разных целей. В частности, геном бактерий реплицирует ДНК-полимераза III. ДНК-полимераза I и II — это вспомогательные ферменты, участвующие в процессах исправления ошибок в ДНК (репарации). ДНК-полимераз в репликативной вилке должно быть две, поскольку цепочек ДНК тоже две.
Необходимо помнить, что полимераза может синтезировать ДНК только в одном направлении от 5’- к 3’- концу. Эти штрихи берутся от нумерации атомов в сахаре. Дезоксирибоза — пятиатомный сахар. Она является необходимым звеном, образующим сахарофосфатный остов. В образовании этого остова играют роль 3 и 5 атомы углерода. Соответственно, у сахара, у сахарофосфатного остова, у ДНК и РНК есть 3’-конец и 5’- конец. Синтезироваться молекула ДНК может только путем присоединения новых нуклеотидов к растущему 3’-концу. Энергия для синтеза ДНК берется от нуклеотид-3-фосфатов, которые одновременно являются и структурными элементами ДНК. При присоединении нуклеотида отщепляется пирофосфат, который тут же разрушается специальным ферментом (пирофосфатазой) еще на две отдельных молекулы ортофосфорной кислоты, тем самым, делая эту реакцию необратимой.
Почему ДНК-полимераза присоединяет нуклеотиды к 3’-концу молекулы? Может быть, это связано с тем, что при таком синтезе ДНК-полимераза может репарировать собственные ошибки. Если фермент присоединил неправильный нуклеотид, а потом «осознал», что ошибся, то он может просто сделать шаг назад, удалить нуклеотид и сделать все, как было. Если бы синтез шел наоборот, так нельзя было бы сделать. На самом деле это точно не известно и причины могут быть другими.
Поскольку нить ДНК растет в одном направлении, и ферментный комплекс движется в одном направлении, как осуществляется синтез комплементарной цепи?
Дело в том, что вторая цепь ДНК образует петлю и разворачивается так, чтобы обе ДНК-полимеразы в составе ферментного комплекса могли двигаться вместе (рис. 3). В результате, на второй цепи ДНК синтез идет кусочками, с перерывами. Эта цепь называется отстающей, другая — лидирующей. Время от времени фермент перескакивает на какое-то расстояние по цепи ДНК дальше и синтезирует дочернюю цепь до следующего перескакивания. Синтез с отстающей цепи происходит фрагментами. Эти фрагменты названы в честь японского ученого, который их описал, — фрагментами Оказаки. Фрагменты Оказаки сшиваются лигазой.
Понятие о репликоне и репликаторе
Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. ген-репликатор. replicator - репликатор, ген-репликатор.
Генетический фактор, контролирующий репликацию участка ДНК (группы генов), т.е. репликона; Р.
функционально связан с геном-инициатором, а в отсутствии репликации Р. репрессирован.
Хромосомная теория наследственности
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения хромосомной теории наследственности:
• Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
• Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
• Гены расположены в хромосоме в линейной последовательности.
• Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола).
• Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
• Каждый биологический вид характеризуется определенным набором
