В начале своего развития тотипотентная зигота содержит, доставшиеся от яйцеклетки, все ключевые транскрипционные и эпигенетические факторы, регулирующие процессы развития. Переключение зиготической программы развития на эмбриональную происходит на уровне поздней зиготы и двухклеточной стадии, когда начинается транскрипция. За этим следует предимплантационное развитие, в процессе которого происходит около 6 делений дробления с образованием бластоцисты – уникальная стадия развития
у млекопитающих, не имеющая аналогов у других организмов. Бластоциста состоит из ~ 60 клеток, из которых часть составляет ВКМ, представленных плюрипотентными клетками примитивной эктодермы, и специализированные клетки внешней трофэктодермы, которые необходимы для имплантации и развития плаценты. ВКМ является основой для всех соматических тканей и половых клеток во взрослом организме. После имплантации ВКМ начинает формировать клетки эпибластов, которые обладают плюрипотентностью, о чем говорит экспрессия гена Oct4. Эти клетки отвечают на сигналы, исходящие от экстраэмбриональных тканей, которые направляют дифференцировку и инициируют гаструляцию.
Одно из наиболее ранних событий в развитии – появление первичных половых клеток в начале гаструляции. Они являются высоко специализированными, с установленной специфической программой транскрипции, которая не запрещает реализацию судьбы соматической клетки. Важно, что только в этих клетках экспрессируются гены, поддерживающие плюрипотентность после гаструляции. Транскрипционная программа, используемая этими клетками, должна так же регулировать эпигенетическое репрограммирование генома, включая полногеномное стирание имеющихся эпигенетических модификаций, характерных для этой группы клеток и являющихся необходимым шагом к конечному тотипотентному состоянию.
ВКМ и примордиальные герминальные клетки (ПГК) в свою очередь являются предшественниками плюрипотентных эмбриональных стволовых (ЭСК) и эмбриональных герминальных (ЭГК) клеток, которые могут быть получены и поддерживаться в культуре in vitro. Недавно, плюрипотентные стволовые клетки были получены из стволовых клеток сперматогониев. Это позволяет предполагать, что транкрипционная сеть и эпигенетические регуляторы, способные поддерживать плюрипотентность, возникли в процессе раннего развития половых клеток. ЭСК демонстрируют постоянное плюрипотентное состояние in vitro, но только похожее на преходящее плюрипотентное состояние клеток примитивной эктодермы in vivo. Для создания и поддержания ЭСК необходимы специфические цитокины. Фактор ингибирования лейкемии (LIF) и белок морфогенеза костной ткани (BMP4) являются ключевыми факторами, которые не только модифицируют клетки примитивной эктодермы и вызывают необходимые реакции в процессе создания ЭСК, но и поддерживают плюрипотентность в культуре. Если эти цитокины не добавлять в культуру, то ЭСК начинают дифференцироваться, подобно клеткам примитивной эктодермы. Это подчеркивает временную природу плюрипотентности клеток примитивной эктодермы, т.к. они очень быстро переходят в другую стадию развития in vivo, но могут поддерживаться бесконечно долго как ЭСК in vitro. Поскольку ЭСК и ЭПК не имеют эквивалента in vivo, они представляют собой уникальное эпигенетическое состояние (Рис. 14).
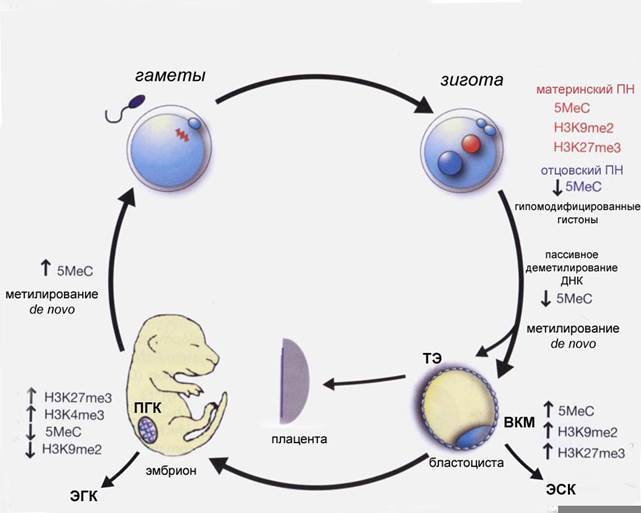
Рис. 14. Генетическая и эпигенетическая регуляция плюрипотентности на ранних этапах эмбрионального развития мыши. ПН- пронуклеус; ВКМ – внутренняя клеточная масса; ТЭ – трофэктодерма; ЭСК – эмбриональные стволовые клетки; ЭГК – эмбриональные герминальные клетки; ПГК – примордиальные герминальные клетки;
2.2. От тотипотентности к плюрипотентности.
2.2.1. Зигота: создание базы для транскрипции.
Во время оплодотворения, когда родительские геномы соединяются в цитоплазме ооцита для формирования тотипотентной зиготы, отцовский геном очень сильно отличается от материнского и должен приобрести подходящее эпигенетическое состояние, чтобы на равных участвовать в развитии (Рис 15).

Рис. 15. Эпигенетические события, происходящие в материнском и отцовском пронуклеусах (а), на стадии бластоцисты в ВКМ и ТЭ в норме (б) и клонированном эмбрионе (с). Зеленый цвет – модификации, разрешающие транскрипцию, красный – запрещающие; F – оплодотворение; PN0-5 – пронуклеарные стадии; S – сингамия.
Первоначально отцовский геном очень плотно упакован с помощью протаминов, которые достаточно быстро замещаются гистонами. Замена происходит до S-фазы, когда определенный вариант гистона H3.3 используется нуклеосомами, т.к. канонический гистон Н3.1 отсутствует в
отцовском пронуклеусе до репликации ДНК. Кроме указанной эпигенетической асимметрии между родительскими геномами, различия касаются модификации гистонов и локализации многочисленных эпигенетических модификаторов, таких как Ezh2.
В отцовском пронуклеусе гистоны модифицированы по-своему. Если H3K4me1, H3K9me1 и H3K27me1 определяются перед оплодотворением, то H3K4me3, H3K9me2 и H3K27me3 обнаруживаются только после репликации. Кроме того, происходит быстрое и почти полное деметилирование ДНК. Молекулярный механизм глобального деметилирования ДНК неизвестен, но правильная эпигенетическая конфигурация отцовского хроматина совершенно необходима, потому что в материнском геноме подобных модификаций не происходит. Т.к. метилирование гистонов может вызывать метилирование
ДНК, по-крайней мере, в определенных участках генома, различия в модификации гистонов между родительскими пронуклеусами можно объяснить защитой материнского генома от претерпеваемого деметилирования ДНК.
Было показано, что PGC7/Stella необходим для предотвращения деметилирования ДНК материнского генома; в ооцитах, дефектных по этому гену, происходит глобальное деметилирование материнского генома. Но, поскольку PGC7 обнаружен как в материнском, так и отцовском пронуклеусах, видимо, существуют и другие факторы, которые защищают материнский геном от деметилирования ДНК.