Глюконеогенез из пировиноградной кислоты
Глюконеогене́з — метаболический путь, приводящий к образованию глюкозы из неуглеводных соединений (в частности, пирувата). Наряду с гликогенолизом, этот путь поддерживает в крови уровень глюкозы, необходимый для работы многих тканей и органов, в первую очередь, нервной ткани и эритроцитов.
Глюконеогенез протекает в основном в печени, но менее интенсивно он протекает также в корковом веществе почек и слизистой кишечника.
После того как в мыщцах истощается запас гликогена, основным источником пирувата становятся аминокислоты, образующиеся после деградации белков. При этом более 30% аминокислот, поступающих из крови в печень, приходится на аланин — одну из гликогенных аминокислот, углеродный скелет которой используется в печени как предшественник для синтеза глюкозы.. Другим источником пирувата является лактат, который накапливается в интенсивно работающих мышцах в процессе анаэробного гликолиза, когда митохондрии не успевают окислить накапливающийся НАДН. Лактат транспортируется в печень, где снова превращается в пируват, а затем в глюкозу и гликоген.

Суммарное уравнение биосинтетических реакций глюконеогенеза, приводящих к образованию глюкозы из пирувата, выглядит так:
2 Пируват + 4ATP + 2GTP + 2NADH + 2H+ + 4H2O → глюкоза + 4ADP + 2GDP + 6Pi + 2NAD+.
Ниже рассмотрены 3 стадии глюконеогенеза, отличные от реакций гликолиза, проведённых в обратном направлении.
Образование фосфоенолпирувата из пирувата(1 стадия)
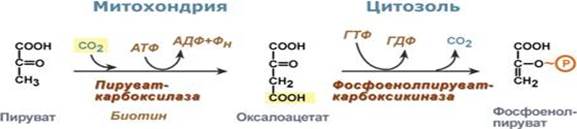
Первой реакцией глюконеогенеза является превращение пирувата в фосфоенолпируват (ФЕП). Фосфорилирование пирувата осуществляется «окольным путём», для реакций которого у эукариот необходимы и цитозольные, и митохондриальные ферменты.
Первоначально пируват под влиянием пируваткарбоксилазы и при участии СО2 и АТФ карбоксилируется с образованием оксалацетата:
Пируват + НСО3- + ATP → оксалоацетат + ADP + Pi
В этой реакции карбоксилирования участвует биотин как переносчик активированного бикарбоната. Так как в митохондриальной мембране отсутствуют белки-переносчики оксалоацетата, до экспорта в цитозоль образовавшийся при пируваткарбоксилазной реакции оксалоацетат должен быть обратимо восстановлен в малат митохондриальным ферментом малатдегидрогеназой с затратой NADH:
Оксалоацетат + NADH + H+ ⇌ L-малат + NAD+.
Митохондриальная малатдегидрогеназа участвует и в глюконеогенезе, и в цикле трикарбоновых кислот, осуществляя и прямую, и обратную реакции. Оксалоацетат может также переноситься из митохондрии в цитозоль после трансаминирования в аспартат.
Малат покидает митохондрию через специальный белок-транспортер на внутренней митохондриальной мембране, и в цитозоле он вновь окисляется до оксалоацетата с образованием цитозольного NADH:
Малат + NAD+ → оксалоацетат + NADH + H+[10].
После этого оксалоацетат превращается в фосфоенолпируват под действием фосфоенолпируваткарбоксикиназы. В этой Mg2+-зависимой реакции донором фосфорильной группы выступает GTP:
Оксалоацетат + GTP ↔ Фосфоенолпируват + СО2 + GDP.
Образование фруктозо-6-фосфата из фруктозо-1,6-бифосфата(2 стадия)
Фосфоенолпируват, образовавшийся из пирувата, в результате ряда обратимых реакций гликолиза превращается во фруктозо-1,6-бифосфат.
Второй реакцией гликолиза, которая не может дублироваться обратной реакцией в глюконеогенезе, является фосфорилирование фруктозо-6-фосфата фосфофруктокиназой-1.
Фруктозо-1,6-бифосфат + Н2О → фруктозо-6-фосфат + P
Данная реакция катализируется специфической фосфатазой.
Образование глюкозы из глюкозо-6-фосфата(3 стадия)
Третья реакция является последней реакцией глюконеогенеза: дефосфорилирование глюкозо-6-фосфата с образованием глюкозы. Если бы гексокиназа осуществляла эту обратную реакцию, то ей бы сопутствовал перенос фосфорильной группы с глюкозо-6-фосфата на ADP с образованием ATP, что энергетически невыгодно. Реакция, катализируемая глюкозо-6-фосфатазой, не включает синтеза ATP и представляет собой простой гидролиз фосфатного эфира:
Глюкозо-6-фосфат + Н2 → глюкоза + Pi.
Глюкоза, образовавшаяся в ходе глюконеогенеза в печени и почках или поглощённая с пищей разносится по кровотоку к этим тканям, в числе которых — мозг и мышцы.

Глюконеогенез – это синтез глюкозы из неуглеводных компонентов: лактата, пирувата, глицерола, кетокислот цикла Кребса и других кетокислот, из аминокислот. Все аминокислоты, кроме кетогенных лейцина и лизина, способны участвовать в синтезе глюкозы. Углеродные атомы некоторых из них (глюкогенных) полностью включаются в молекулу глюкозы, некоторых (смешанных) частично.
Кроме получения глюкозы, глюконеогенез обеспечивает и уборку лактата, постоянно образуемого в эритроцитах или при мышечной работе, и глицерола, являющегося продуктом липолиза в жировой ткани.
Как известно, в гликолизе существуют три необратимые реакции: пируваткиназная (десятая), фосфофруктокиназная (третья) и гексокиназная (первая). В этих реакциях происходит высвобождение энергии для синтеза АТФ. Поэтому в обратном процессе возникают энергетические барьеры, которые клетка обходит с помощью дополнительных реакций.
Глюконеогенез включает все обратимые реакции гликолиза, и особые обходные пути, т.е. он не полностью повторяет реакции окисления глюкозы. Его реакции способны идти во всех тканях, кроме последней глюкозо-6-фосфатазной реакции, которая идет только в печени и почках. Поэтому, строго говоря, глюконеогенез идет только в этих двух органах.
Обход десятой реакции гликолиза
На этом этапе глюконеогенеза работают два ключевых фермента – в митохондриях пируваткарбоксилаза и в цитозоле фосфоенолпируват-карбоксикиназа, при этом в реакциях тратятся два макроэрга - АТФ и ГТФ.
В химическом плане обходной путь десятой реакции выглядит достаточно просто:
Упрощенный вариант написания обхода десятой реакции гликолиза
Однако дело в том, что пируваткарбоксилаза находится в митохондрии, а фосфоенолпируват-карбоксикиназа – в цитозоле. Дополняет проблему непроницаемость митохондриальной мембраны для оксалоацетата. Зато через мембрану может пройти малат, предшественник оксалоацетата по ЦТК.
Поэтому в реальности все выглядит более сложно:
1. В цитозоле пировиноградная кислота может появиться при окислении молочной кислоты и в реакции трансаминирования аланина. После этого пируват симпортом с ионами Н+, движущимися по протонному градиенту, проникает в митохондрии. В митохондриях пируваткарбоксилаза превращает пировиноградную кислоту в оксалоацетат.
Пируваткарбоксилазная реакция идет в клетке постоянно, так как оксалоацетат является главным регулятором скорости ЦТК. Реакция называется анаплеротической (пополняющей) реакцией ЦТК.
2. Далее оксалоацетат мог бы превратиться в фосфоенолпируват, но для этого сначала он должен попасть в цитозоль. Поэтому происходит реакция восстановления оксалоацетата в малат при участии малатдегидрогеназы. В результате малат накапливается, выходит в цитозоль и здесь превращается обратно в оксалоацетат.
Повернуть малатдегидрогеназную реакцию ЦТК вспять позволяет избыток НАДН в митохондриях. НАДН поступает из β-окисления жирных кислот, активируемого в гепатоците при голодании.
3. В цитоплазме фосфоенолпируват-карбоксикиназа осуществляет превращение оксалоацетата в фосфоенолпируват, для реакции требуется энергия ГТФ. От молекулы отщепляется тот же углерод, что и присоединяется.
