Моховидные представляют совершенно особую линию развития высших растений. Главная отличительная особенность этой группы состоит в том, что в жизненном цикле моховидных преобладает гаметофитная стадия или, как еще иначе говорят, гаметофитное поколение, хотя этот термин и не совсем удачен. Спорофит же в значительной степени редуцирован и развивается на гаметофите. Гаметофитная стадия — это этап жизненного цикла растений от споры до зиготы. На гаметофитах образуются мужские и женские гаметангии, т. е. половые органы, называемые соответственно антеридиями и архегониями, таким образом, это половая, гаплоидная стадия, в противоположность спорофиту. Спорофитная стадия есть этап жизненного цикла от момента образования зиготы до начала спорообразонания. На спорофитах развиваются спорангии, в которых образуются споры; это бесполая, диплоидная стадия индивидуального развития. Моховидные — второй по численности после цветковых отдел высших растений, насчитывающий около 25 тыс. видов, встречающихся практически на всех континентах, в самых разнообразных, порой экстремальных, условиях обитания. Особенно широко эта группа растений распространена во влажных местообитаниях, что связано с их биологическими особенностями. Они встречаются в лесах умеренного пояса и тропиков, на болотах, в тундрах, во влажных горных лесах, по берегам ручьев и лесных проток. Среди них есть и виды, живущие в пресноводных водоемах. Морских обитателей нет, хотя некоторые виды поселяются на скалах в полосе прибоя. Лишь относительно небольшая часть моховидных встречается в сухих местообитаниях, таких, как степи, саванны и даже каменистые пустыни, сохраняя при этом жизнеспособность в течение нескольких лет отсутствия влаги. Эти растения обитают на самых разнообразных субстратах: почве, скалах, камнях, стволах деревьев, валежнике, образуя при этом во влажных лесах мощные моховые подушки, впитывающие, в силу гигроскопичности, большие количества воды. В тропических лесах некоторые виды мхов могут поселяться на поверхности листовых пластинок других высших растений, например, цветковых или папоротникообразных и образуют особую группу так называемых эпифиллов («эпи» — на и «филлос» лист), наносящих растению-хозяину значительный вред, так как они располагаются на активно ассимилирующих участках листовой поверхности. Вместе с тем, настоящих паразитов среди моховидных нет. Большинство моховидных являются тенелюбивыми п встречаются там, где из-за затенения не могут произрастать другие растения. Поэтому в темных хвойных лесах развивается мощный моховой покров. Моховидные имеют совершенно особое положение в природе, поселяясь на таких субстратах и в таких местообитаниях, которые являются совершенно недоступными для других групп растений. Являясь важнейшими продуцентами, они играют большую роль на начальных стадиях почвообразовательных процессов, регулируют водный баланс суши в природе. При этом моховидные совершенно не выносят антропогенного загрязнения среды, особенно серосодержащими соединениями, хотя некоторые виды мхов поселяются в крупных городах, на территориях промышленных предприятий, у автомобильных дорог и пр. Хозяйственное значение мхов для человека невелико. Домашними животными они практически не поедаются. С давних пор хорошо известны антисептические свойства сфагновых мхов (сфагнума), которые обладают еще и значительной гигроскопичностью. Все это позволяло применять эти растения как перевязочный материал. Велика роль мхов как торфообразователей. Торф же применяют в качестве топлива, для изготовления теплоизолирующих материалов, и т. д. Омоховение земель вызывает их переувлажнение и как следствие этого — заболачивание. Вместе с тем, наличие мохового покрова является одним из основных стабилизирующих факторов в условиях вечной мерзлоты.
Морфология моховидных
Гаметофиты моховидных — это, как правило, низкорослые растения, редко достигающие несколько десятков сантиметров в длину (как, например, водные мхи). Все структуры, развивающиеся на гаметофите, у моховидных являются аналогичными (но не гомологичными) соответствующим структурам других высших растений, в жизненном цикле которых преобладает спорофитная стадия. У моховидных нет настоящих корней, а их функции выполняют ризоиды. Что касается других вегетативных органов, то настоящие стебли и листья, характерные для спорофита остальных высших растений, у моховидных также отсутствуют. Однако на их гаметофите развиваются листовидные и стеблевидные структуры, которые еще иначе называют филлидиями и каул лидиями соответственно. Анатомическая организация мхов также проста. Обитание в относительно постоянных условиях (благодаря своей низкорослости они поселяются в экологически наиболее стабильном припочвенном или присубстратном слое), приводит к тому, что дифференциация тканей выражена слабо. В гаметофите моховидных, достаточно условно, можно выделить ассимиляционную, механическую, покровную и проводящую ткани, и то только у наи-более эволюционно продвинутых групп. Настоящих устьиц нет — вместо них имеются поры, лишенные замыкающих клеток. Гаметангии, или органы полового размножения моховидных, имеют весьма сложное строение. Мужские органы полового размножения — антеридии — представляют собой однослойные мешочкоподобные образования на ножке, заполненные сперматозоидными клетками. Из каждой сперматозоидной клетки образуется двужгутиковый сперматозоид. Архегонии — женские половые органы бутылеобразные структуры, состоящие из брюшка, в котором помещается яйцеклетка, шейки и ножки. Как архегонии, так и антеридии могут быть либо непосредственно погружены в ткань гаметофита, либо возвышаться над ним на особых подставках. Среди моховидных можно встретить как однодомные, так и двудомные виды. У однодомных и архегонии, и антеридии развиваются на одном и том же растении, у двудомных же — на разных, и в связи с этим выделяют мужские и женские особи.

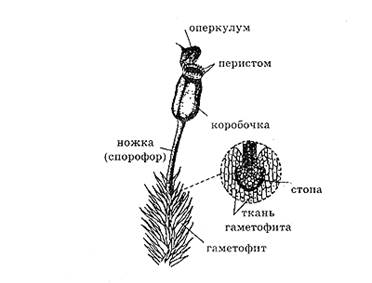
Спорофит, или бесполая стадия жизненного цикла, имеет более простое строение, чем у других групп высших растений. Он не способен укореняться и располагается на гаметофите. Спорофит моховидных называют иначе — спорогоном. Он, как правило, состоит из трех элементов:
- коробочки (или спорангия), в которой развиваются споры;
- ножки (или спорофора), на которой располагается коробочка;
- стопы, обеспечивающей физиологическую связь с гаметофитом; именно стопа погружена в ткань гаметофита и окружена чехлом из лабиринтных клеток, обладающих способностью накапливать воду. Разрастаясь, архегоний может покрывать коробочку и образовывать колпачок или калиптру. У листостебельных мхов коробочка имеет крышечку или оперкулум, а отверстие коробочки окружают зубцы или перистом, что является приспособлением для вскрытия и рассеивания спор. На спорогоне моховидных имеются настоящие устьица с замыкающими клетками. Внутренняя организация спорофита относительно проста. На поперечном срезе можно выделить эпидерму, паренхимную ткань и проводящий тяж, состоящий из гидроидов — мертвых клеток, лишенных протопласта (их функция — транспорт воды и минеральных веществ), и лептоидов — живых клеток, окружающих гидроиды (выполняют функцию транспорта продуктов ассимиляции). Аналогичные образования имеются и в гаметофите (см. выше).

Жизненный цикл моховидных
Как уже говорилось, мужские и женские гаметы развиваются в гаметангиях — органах полового размножения (антеридиях и архегониях), формируясь на гаметофите.
Для того, чтобы сперматозоид достиг яйцеклетки и произошло оплодотворение, совершенно необходима вода, собственно по этой причине моховидные и поселяются в основном во влажных местообитаниях. Попав в водную среду, двужгутиковый сперматозоид перемещается к архегонию и попадает на его шейку. По мере созревания архегония шейковые канальцевые клетки разрушаются и образуется канадец, по которому сперматозоид проникает в брюшко, где происходит оплодотворение яйцеклетки и образование зиготы. После этого зигота делится, и внутри архегония развивается зародыш, который как и зигота является диплоидным образованием, в противоположность гаметам, гаметангиям и всем структурам гаметофита, которые являются гаплоидными. Затем зародыш растет, дифференцируется и в конце концов образуется спорогон, имеющий типичное для той или иной группы моховидных строение. На ранних стадиях развития спорогон полностью зависит от гаметофита и получает от него необходимые питательные вещества. После этого наступает период, когда спорогон приобретает способность фотосинтезироать и становится относительно независимым от гаметофита. И, наконец, на заключительном этапе онтогенеза спорогона, к моменту созревания спор, он теряет фотосинтезирующую активность и становится вновь зависимым от гаметофита. Внутреннее содержание коробочки созревающего спорогона представлено спорогенной тканью — археспорием. Первично клетки археспория диплоидны, но по мере созревания каждая клетка подвергается мейотическому редукционному делению и дает начало четырем гаплоидным клеткам спор. Кроме спор, в спорангиях печеночных мхов формируются особые клетки — элатеры (пружинки), функция которых — разрыхление споровой массы и выброс спор из спорангия. Так или иначе, высвободившиеся споры, попав в благоприятные условия на подходящий субстрат, прорастают. При прорастании спор образуется протонема — юношеская стадия в развитии гаметофита, морфологически отличная от «взрослого» растения. Протонема может быть либо пластинчатой, либо нитчатой, похожей на нить зеленых водорослей, и обладает способностью к активному фотосинтезу. По мере дальнейшего развития формируются зрелые гаметофиты, на которых развиваются гаметангии, и цикл таким образом замыкается.
Систематика моховидных
Отдел моховидных принадлежит царству растений и делится по современным представлениям на три класса: антоцеротовые, печеночные и бриевые (или листосте-бельные) мхи.
Антоцеротовые мхи. В настоящее время насчитывается около 300 видов антоцеротовых мхов. Распространены они, главным образом, в тропических и умеренно теплых странах, предпочитая влажные затененные места. Гаметофит антоцеротовых представлен слоевищем, состоящим из однородных клеток и сплюснутым в спинобрюшном (дорзовентральном) направлении. Каждая клетка слоевища содержит один крупный хлоропласт водорослевого типа — хроматофор. На нижней стороне слоевища имеются полости (вмятины), заполненные слизью. В этой слизи обычно обитают симбионты — сине-зеленые водоросли. Гаметангии развиваются в ткани слоевища. Основной отличительной особенностью этой группы является то, что у основания спорогона располагается слой меристематической ткани, благодаря чему он может расти на протяжении всего своего существования от основания вверх. Представители: антоцерос гладкий, нототилас округлый и др. Печеночные мхи насчитывают около 8500 видов из 280 родов. Гаметофит представлен слоевищем, которое может дихотомически, т. е. вильчато ветвиться, хотя имеются и облиственные печеночники. На нижней стороне слоевища находятся одноклеточные ризоиды, а также полости, аналогичные полостям антоцеротовых мхов, но заполненные воздухом, так называемые воздушные камеры. На талломе, кроме того, образуются выводковые почки или геммы в особых выводковых корзинках. Из таких почек могут развиваться новые гаметофиты, генетически идентичные материнскому растению, таким образом, это один из способов бесполого размножения. Органы полового размножения (гаметангии) располагаются в тканях особых подставок (гаметофоров), поднимающихся над поверхностью слоевища на ножках. Гаметофиты многих печеночных мхов (например, маршанции) — однополы: либо женские, либо мужские. Спорофит состоит из трех частей: спорангия, ножки и стопы. Коробочка (спорангий) просто растрескивается на ряд лепестковидных сегментов. В споровой массе располагаются одноклеточные элатеры, обладающие большой гигроскопичностью и способностью скручиваться и раскручиваться. Протонема у печеночных слабо развита. К печеночным мхам относятся представители родов маршанция,риччия и др. Листостебельные мхи — самая большая группа моховидных, насчитывающая около 14500 (до 18000) видов из более чем 700 родов. Гаметофит представлен стеблевидным образованием — каулидием, который может обладать либо вертикальным ростом, либо полегать. На каулидии располагаются филлидии — листоподобные сидячие, поперечно прикрепленные образования, состоящие из одного слоя клеток. Внутренняя организация имеет выраженные элементы тканевой дифференцировки. Ризоиды — многоклеточные, что отличает эту группу от печеночников. Органы полового размножения — архегонии и антеридии — располагаются либо на верхушке основного «побега» — каулидия, либо (если он ветвится) — на концах боковых веточек. Причем гаметангии, как правило, собраны в группы, окруженные специализированными покровными <. листьями» — фил-лидиями. Спорофит относительно сложно устроен: стенка спорангия — многослойная, на начальных этапах онтогенеза — фотосинтезирует, имеет настоящие устьица, на поперечном срезе ножки выражены проводящие ткани (см. выше). Внутри спорангия располагается колонка, представленная бесплодной (стерильной) тканью. Спорангий дифференцируется на собственно коробочку и крышечку, кроме того, развит перистом (см. выше). Элатеры у листостебельных мхов не образуются. Протонема, как правило, сложно устроена, может быть либо. нитчатой, либо пластинчатой. В системе листостебельных мхов выделяют три подкласса: андреевые, сфагновые и собственно настоящие (или бриевые) мхи. Представители: р.р. сфагнум (эти мхи являются важнейшими торфообразователями), кукушкин юн, бриум, мниум и др.