Глава 8. Физиология синаптической передачи.
Термин и понятие «синапс», «синаптическая передача» был введен в физиологию Ч. Шеррингтоном в 1897 году. Исследуя деятельность ЦНС, он предположил, что между собой нейроны сообщаются с помощью специального – так называемого синаптического механизма. Последующие годы подтвердили его предположение. В разработке учения о синапсах большую роль сыграли О. Леви, Г. Дейл, Д. Ленгли, Дж. Экклс, Дель-Кастильо, В. Катц, а также многие отечественные физиологи – А.Ф. Самойлов, А.В. Кибяков, Х.С. Коштоянц, Д.Г. Магазанник, Г.И. Полетаев, Е.Е. Никольский, А.Л. Зефиров, Р.А. Гиниатуллин и другие.
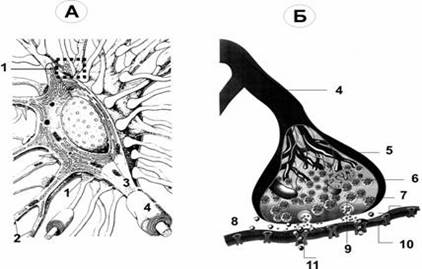
Рис. Межнейронный (аксосоматический) синапс. А. На поверхности перикариона заканчивается множество (например, на мотонейронах до 10 000) концевых разветвлений (пресинаптических терминалей) аксонов от других нейронов; это аксосоматические синапсы. Один из них (1) представлен в правой части схемы. 2 — ветвления одного из дендритов. 3 — место отхождения аксона (аксонный холмик). 4 — миелинизированный аксон. Б. Между концевой терминалью аксона (5) пресинаптического нейрона и поверхностью перикариона постсинаптического нейрона (10) находится синаптическая щель (8) с диффундирующими в ней молекулами нейромедиатора (9). Для терминали аксона (пресинаптическая часть синапса, 5) характерны митохондрии (6) и содержащие молекулы нейромедиатора синаптические пузырьки (7). В постсинаптическую мембрану вмонтированы рецепторы для связывания нейромедиатора и многочисленные ионные каналы (11). Помимо аксосоматических, между нейронами образуются аксоаксональные, аксодендритические и дендродендритические синапсы. Большинство межнейронных синапсов относится к аксодендритическим (например, в коре больших полушарий — до 98%).
Общая физиология синапса
Классификация синапсов. Синапс – это морфофункцнональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку). Все синапсы ЦНС можно классифицировать следующим образом.
1. По локализации – центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейросекреторный, синапс вегетативной нервной системы). Центральные синапсы можно в свою очередь разделить на аксо-аксональные, аксо-дендритические (дендритные), аксо-соматические, дендро-дендритические, дендро-соматические и т.п. Согласно Г. Шеперду, различают реципрокные синапсы, последовательные синапсы и синаптические гломерулы (различным способом соединенные через синапсы клетки).
2. По развитию в онтогенезе – стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития.
3. По конечному эффекту – тормозные (тормозящие) и возбуждающие.
4. По механизму передачи сигнала – электрические, химические, смешанные.
Химические синапсы можно классифицировать:
а) по форме контакта – терминальные (колбообразные соединения) и преходящие (варикозные расширения аксона);
б) по природе медиатора – холинергические (медиатор – ацетилхолин), адренергические (норадреналин, в отдельных случаях адреналин), дофамннергические (дофамин), серотнинергические (серотонин), ГАМК-ергические (медиатор – гамма-аминомасляная кислота), глицинергические (глицин), глютаматергические (глютамат), пептидергические (медиатор – пептиды, например, вещество Р), пуринергические (медиатор – АТФ), азотергические (медиатор – оксид азота NO) и другие.
Электрические синапсы (эфапсы). Вопрос о них во многом не ясен. До настоящего времени недостаточно четко дифференцируются такие понятия как «электрический синапс» (в ЦНС) и «нексус» (в гладких мышцах, в миокарде). В настоящее время признают, что в ЦНС имеются электрические синапсы. В основном они локализованы в стволе мозга, в том числе они обнаружены между нейронами мезенцефального ядра тройничного нерва, а также между нейронами вестибулярного ядра Дейтерса и ядер нижней оливы в продолговатом мозге. С точки зрения морфологии электрический синапс представляет собой щелевидное образование (размеры щели до 2 нм) с ионными мостиками-каналами между двумя контактирующими клетками. Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно проходят через такой щелевидный контакт и возбуждают, т.е. индуцируют генерацию ПД в соседней клетке. В целом, эфапсы обеспечивают очень быструю передачу возбуждения. Но в то же время с помощью этих синапсов нельзя обеспечить одностороннее проведение, т.к. большая часть таких синапсов обладает двусторонней проводимостью. Кроме того, с их помощью нельзя вызывать торможение эффекторной клетки.
Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса.
Общие представления о строении и механизмах функционирования химического синапса. Химические синапсы (далее по тексту синапсы) представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы).
Химический синапс состоит из трех компонентов (или элементов): пресинаптической части, постсинаптической части и синаптической щели. В пресинаптической части содержится медиатор (трансмиттер), который под влиянием нервного импульса выделяется в синаптическую щель и, связываясь с рецепторами в постсинаптической части, вызывает ряд физиологических эффектов, в том числе изменение ионной проницаемости постиснаптической мембраны, что приводит к ее деполяризации (в возбуждающих синапсах) или гиперполяризации (в тормозных синапсах).
Пресинаптическая часть (или пресинаптический элемент, пресинпас) образуется аксоном по его ходу (проходящий синапс) или представляет собой расширенную конечную часть аксона (концевой бутон). В ней содержатся митохондрии, агранулярная эндоплазматическая сеть, нейрофиламенты, нейротрубочки и синаптические пузырьки диаметром 20-65 нм, в которых находится нейромедиатор. Форма и характер содержимого пузырьков зависят от находящихся в них нейромедиаторов. Круглые светлые пузырьки обычно содержат ацетилхолин, пузырьки с компактным плотным центром – норадреналин, крупные плотные пузырьки со светлым подмембранным ободком – пептиды. Медиаторы вырабатываются в теле нейрона и механизмом быстрого аксонного транспорта переносятся в окончания аксона, где происходит их депонирование. Частично синаптические пузырьки образуются в самом синапсе путем отщепления от цистерн агранулярной эндоплазматической сети. На внутренней стороне плазмолеммы, обращенной к синаптической щели, т.е. пресинаптической мембраны имеется пресинаптическое уплотнение, образованное фибриллярной гексагональной белковой сетью, ячейки которой способствуют равномерному распрелению синаптических пузырьков по поверхности мембраны.
Постсинаптическая часть (или постсинаптический элемент) представлена постсинаптической мембраной, содержащей особые комплексы интегральных белков – синаптические рецепторы (мембранные рецепторы), связывающиеся с нейромедиатором. Мембрана утолщена за счет скопления под ней плотного филаментозного белкового материала (постсинаптическое уплотнение). По обе стороны от постсинаптической мембраны располгаются внестинаптические области, которые играют исключительно важную роль в процессах дальнейшего проведения возбуждения от постиснаптической мембраны.
Синаптическая щель – это еще один компонент синапса. Ее ширина обычно варьирут от 20-30 нм до 50 нм. Во многих синапсах синаптическая щель содержит поперчно расположенные гликопротеиновые интрасинаптические филаменты толщиной 5 нм, которые являются элементами специализированного гликокаликса. За счет этих элементов обеспечивается адгезивные связи пре- и постсинаптических частей, а также направленная диффузия медиатора.
В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка – это синапс между нейронами, концевая пластинка – это постсинаптическая мембрана мионеврального синапса, моторная бляшка – это пресинаптическое окончание аксона на мышечном волокне.
Любой химический синапс, независимо от природы медиатора и хеморецептора, активируется под влиянием потенциала действия, распространяющегося к пресинапсу от тела нейрона. Под влиянием потенциала действия происходит деполяризация пресинаптической мембраны, что повышает проницаемость кальциевых каналов пресинаптической мембраны и приводит к увеличению входа в пресинапс ионов Са2+. В ответ на это происходит высвобождение (выход из пресинапса) 100-200 порций, или квантов, медиатора, что осуществляется путем экзоцитоза. Выйдя в синаптическую щель, медиатор взаимодействует со специфическими рецепторами постсинаптической мембраны. Во многих синапсах основная масса рецепторов – это ионотропные рецепторы. Активированные медиатором они непосредственно регулируют проницаемость ионных каналов постсинаптической мембраны. Кроме того, в ряде синапсов имеются метаботропные рецепторы, при активации которых (за счет вторичных посредников и протеинкиназ) меняется активность внутриклеточных белков-эффекторов, в том числе ионных каналов и ионных насосов. В целом, взаимодействие медиатора с постсинаптическими рецепторами изменяет ионную проницаемость. В синапсах, в которых осуществляется возбуждение постсинаптической структуры, обычно происходит повышение проницаемости для ионов Na+ или Cа2+, что вызывает деполяризацию постсинаптической мембраны. Эта деполяризация получила название – возбуждающий постсинаптический потенциал, или ВПСП (в нервно-мышечном синапсе – потенциал концевой пластинки, или ПКП). Если его величина достигает критического уровня деполяризации, то во внесинаптических областях генерируется ПД. В тормозных синапсах в результате взаимодействия медиатора с рецепторами, наоборот, происходит гиперполяризация (за счет, например, увеличения проницаемости для ионов калия и хлора). Этот вид изменения мембранного потенциала получил название тормозной постсинаптический потенциал, или ТПСП. В гиперполяризованном состоянии клетка снижает свою возбудимость и благодаря этому прекращает отвечать на внешние раздражители или (если она обладала свойством автоматии) уменьшает спонтанную активность.
Одновременно, выделившийся в синаптическую щель медиатор может взаимодействовать с рецепторами, расположенными на пресинаптической мембране. Таким способом регулируется интенсивность последующего высвобождения медиатора, т.е. процесс экзоцитоза. Это получило название антидромного эффекта или явления обратной связи.
После каждого цикла проведения нервного импульса медиатор разрушается и с участием специфического фермента подвергается удалению. Например, ацетилхолин разрушается ацетилхолин-эстеразой, норадреналин – моноаминоксидазой (МАО) и катехол-0-метилтрансферазой (КОМТ). Одновременно происходит обратный захват медиатора (например, норадреналина) или продуктов его расщепления (например, холина при разрушении ацетилхолина) в пресинаптическую структуру (это называется нейрональный захват) либо в постсинаптическую структуру (экстранейрональный захват). Кроме того, снижение концентрации медиатора в синаптической щели достигается путем его простой диффузии во внесинаптические пространства.
Рассмотрим более подробно процессы, происходящие в химическом синапсе.
Синтез медиатора совершается в пресинаптическом элементе, куда из крови или спинномозговой жидкости попадают исходные продукты (предшественники медиаторов) и ферменты, необходимые для его синтеза. Ферменты образуются в соме нейрона и по аксону, примерно со скоростью 6 мм/сутки, транспортируются в пресинаптическое окончание аксона, где используются в процессе синтеза медиатора. Угнетение активности этих ферментов фармакологическим путем может привести к истощению запасов медиатора в синапсе и, следовательно, к снижению его функциональной способности. Затем образовавшийся медиатор путем активного транспорта вводится в синаптические везикулы (мелкие везикулы).
Синтез нейропептидов происходит подобно синтезу пептидных гормонов. Первоначально крупные аминокислотные последовательности образуются на рибосомах и помещаются в эндоплазматический ретикулюм. В цистернах аппарата Гольджи осуществляется протеолитический процесс расщепления крупных полипептидов на фрагменты с образованием активных пептидов, которые включаются в отпочковавшиеся крупные везикулы. Разные пептидные фрагменты могут оказаться в различных везикулах, которые транспортируются в нервные окончания нейрона. Синтез и упаковка в везикулы классических медиаторов и нейропептидов в нейроне происходят параллельно. Поэтому из его нервных окончаний освобождаются несколько различных медиаторов.
Синаптические везикулы и их типы. Любой медиатор в пресинаптических нервных окончаниях содержится в везикулах. Существует два основных типа везикул – мелкие и крупные. Мелкие синаптические везикулы имеют диаметр около 50 нм. Они однородны по размерам и содержат классический медиатор. Крупные электронноплотные везикулы имеют диаметр около 100 нм. Они неоднородны по размерам и содержат пептиды, которые помимо медиаторной (передаточной) функции могут модулировать, т.е. изменять, синаптическую передачу.
Все синаптические везикулы образуются в теле нервной клетки из эндоплазматического ретикулюма и цистерн аппарата Гольджи. Затем они транспортируются по аксону в нервные окончания. Крупные везикулы заполняются медиатором (пептидом) непосредственно в соме нейрона. Мелкие везикулы содержат АТФ, ионы, ферменты, а их мембрана имеет протоновые насосы. Именно с участием этих насосов (главным образом, в пресинаптическом окончании) происходит наполнение мелких везикул медиатором, концентрация которого достигает в них высоких значений (около 100 ммоль/л).
Мелкие синаптические везикулы в пресинаптическом окончании формируют два основных пула – пул 1 и пул 2. Пул 1 – это относительно небольшой пул, готовый к немедленному экзоцитозу. Иначе говоря, это запас доступного медиатора, который может освободиться в течение короткого промежутка времени. Пул 1 содержит около 10-50 везикул и при истощении пополняется в течение 5-12 секунд. Вероятность освобождения кванта из пула 1 пропорциональна количеству везикул в пуле. Размер пула является постоянной величиной и определяет эффективность работы синапса.
Пул 2 – это относительно большой пул, везикулы которого не участвуют в секреции, но могут перемещаться по мере необходимости в пул 1. Иначе говоря, пул 2 – это мобилизационный запас медиатора, упакованного в везикулу, который с определенной скоростью может пополнять запас доступного медиатора. Процесс дислокации везикул из пула 2 в пул 1 регулируется внутриклеточной концентрацией ионов кальция.
Пути освобождения медиатора из синапса. В основе освобождения медиатора лежит процесс экзоцитоза, который представляет собой разновидность активного транспорта, предназначенного в живых системах для выделения в окружающую среду гормонов, медиаторов, модуляторов и других веществ.
Существуют два принципиальных механизма освобождения медиатора и всего содержимого везикулы в синаптическую щель. Первый механизм представляет собой классический экзоцитоз, при котором происходит полное слияние везикулы с пресинаптической мембраной. В этом случае все содержимое везикулы (медиатор, АТФ, ионы, ассоциированные белки и ферменты) оказывается в синаптической щели. Второй механизм – это экзоцитоз, протекающий без полного слияния везикулы с пресинаптической мембраной и с частичным освобождением медиатора. Он характеризуется формированием временной поры (канала) в пресинаптической мембране, которая обладает селективностью (поэтому другие ингредиенты, находящиеся в везикуле, не выходят в синаптическую щель). За счет этой поры полость везикулы сообщается с синаптической щелью: при открытии поры (в момент контакта с пресинаптической мембраной) медиатор по градиенту концентрации частично диффундирует в синаптическую щель. Следовательно, везикула при каждом контакте с пресинаптической мембраной теряет только часть своего содержимого. Поэтому при таком способе функционирования она может участвовать в экзоцитозе многократно.
Организация секреции медиатора. Активная зона. Процессы экзоцитоза в синапсе осуществляются в специализированных пресинаптических структурах, которые получили название активных зон. В каждом синапсе число таких зон достигает 1-5. Активная зона имеет длину 1-3 мкм, ширину – 0,1 мкм; от соседней активной зоны она отдалена на расстоянии 1-2 мкм. Каждая активная зона состоит из пресинаптической плотной полоски, около которой рядами расположены кальциевые каналы (белки), два ряда синаптических везикул, а также специализированные белки экзоцитоза (в том числе соединяющие везикулу с пресинаптической мембраной) и элементы цитоскелета, т.е. сеть актиновых филаментов и микротрубочек. Везикулы связываются с элемегнтами цитоскелета и между собой с участием белка (синапсин I).
Элементы цитоскелета удерживают синаптические везикулы в пуле 1 (и пуле 2, т.е. в резервном пуле) и ограничивают их избыточную мобилизацию, регулируют процессы освобождения, обеспечивают характерную пространственную организацию активных зон и рециркулирование мембраны синаптической везикулы после экзоцитоза.
Синаптический эндоцитоз (реинтернализация и рециклизация везикул).После того, как везикула избавляется от своего содержимого, ее мембрана полностью сливается с мембраной нервного окончания, а затем вновь реинтернализируется (т.е. возвращается в пресинаптическое окончание) через стадию «покрытых» молекул. Этот процесс совершается вне места освобождения везикулы. Кроме того, существует еще один вариант эндоцитоза, т.е. возвращения везикулы в пресинапс. Он заключается в том, что при экзоцитозе мембрана везикулы не сливается с пресинаптической мембраной, поэтому после выброса медиатора везикула быстро реинтернализируется в том же самом месте. Оба этих варианта эндоцитоза и рециклизации синаптической везикулы осуществляются одновременно. Полагают, что популяция немедленно готовых к освобождению синаптических везикул и быстро восстанавливающаяся в результате рециклизации по первому пути, отвечает за передачу возбуждения в синапсе при редкой активности, а вторая популяция (в которой везикулы не сливаются) обеспечивает массивное освобождении медиатора.
Функционирование активной зоны. В ней происходят последовательно 5 процессов. 1. Транспорт (мобилизация) везикулы из пула 2 в пул 1 за счет элементов цитоскелета. 2. Стыковка (докирование) везикулы с местом освобождения в активной зоне. 3. Подготовка (прайминг) везикулы к экзоцитозу, которая заключается в комплексной трансформации белкового комплекса экзоцитоза (в результате такой подготовки везикула готова к выбросу своего содержимого и ожидает лишь входа ионов кальция). На стадии прайминга везикулы должны быть предохранены от немедленного слияния и в тоже время они должны быть готовы быстро излить свое содержимое в синаптическую щель в ответ на вход ионов кальция в пресинаптическое окончание. 4. Экзоцитоз – слияние мембраны везикул с плазматической мембраной. Для осуществления этого процесса необходимо открытие потенциалозависимых Са-каналов и увеличение концентрации кальция в области везикулы. 5. Рециклизация везикулы посредством эндоцитоза с последующим заполнением медиатором и включением ее в пулы 1 или 2.
Активные зоны (рис. 6–3). Секреция нейромедиатора осуществляется в специализированных участках пресинаптического нервного окончания — активных зонах — участках утолщения пресинаптической мембраны. Активная зона состоит из «плотной полоски» на пресинаптической мембране и сгруппированных около неё синаптических пузырьков, потенциалозависимых кальциевых каналов, специальных белков экзоцитоза и элементов цитоскелета. Количество активных зон в нервно-мышечном синапсе достигает 30–40, в межнейронных синапсах — около десятка. Активные зоны расположены против скоплений рецепторов в постсинаптической мембране, что уменьшает задержку в передаче сигнала, связанную с диффузией нейромедиатора в синаптической щели.

Рис. 6–3. Активные зоны нервно-мышечного синапса расположены напротив постсинаптических складок — участков скоплений холинорецепторов. Пресинаптическая мембрана слева расщеплена на два листка.
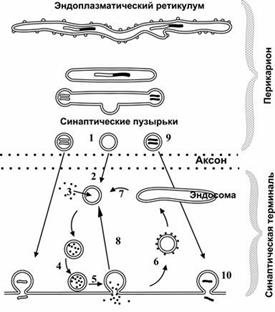
Жизненный цикл синаптических пузырьков (рис. 6–4). Синаптические везикулы образуются в теле нейрона в эндоплазматическом ретикулуме и комплексе Гольджи (1) и с аксонным транспортом поступают в нервные окончания (2). В нервном окончании мелкие синаптические пузырьки посредством активного транспорта заполняются медиатором (3) и передвигаются к пресинаптической мембране (4). Освобождение медиатора (5) может осуществляться посредством экзоцитоза с полным («классический» механизм) либо неполным (механизм «kiss and run») слиянием. Первый вид экзоцитоза сопровождается встраиванием мембраны везикулы в пресинаптическую, опорожнением пузырька, а затем посредством эндоцитоза образуются покрытые клатрином везикулы (6), которые затем проходят стадию эндосомы (7) и снова заполняются медиатором (3). Второй вид экзоцитоза характеризуется образованием временной поры, соединяющей полость пузырька с синаптической щелью. После выделения медиатора везикула не встраивается в пресинаптическую мембрану, а отпочковывается от неё (8) и повторно заполняется медиатором (3). Крупные синаптические везикулы заполняются медиатором в теле клетки (9), их экзоцитоз происходит в других участках пресинаптической мембраны, а эндоцитоз опорожнённых пузырьков отсутствует.

Секретосома. Будучи главным элементом системы секреции медиатора, секретосома представляет собой синаптическую везикулу, докированную в области активной зоны, а также кальциевый канал (каналы) и белки, ответственные за экзоцитоз и эндоцитоз, т.е. белковый комплекс. Область, в которой повышена концентрация ионов кальция, и где происходит экзоцитоз, называется кальциевым микродоменом. Таким образом, секретосомы представляют собой функциональную единицу активной зоны, или пула 1. Количество секретосом в отдельной активной зоне определяется размерами активной зоны (обычно оно равно нескольким десяткам). Каждая секретосома может участвовать в вызванном и спонтанном высвобождении медиатора.
Белковый аппарат экзоцитоза и эндоцитоза. Различают как минимум 8 различных белков, имеющих отношение к процессам экзоцитоза медиатора и эндоцитоза везикулы. 1.Синаптотагмин (65 кД) – белок, связанный с везикулой и предназначенный для связывания ионов кальция (кальциевый сенсор). 2.Синаптобревин (18 кД) – белок, связанный с везикулой и предназначенный для образования комплекса (везикула + пресинаптическая мембрана). 3. Синапсин I (86 кД) – белок, связывающий везикулы с цитоскелетом и тем самым участвующий в процессах мобилизации везикул. 4.Синтаксин (35 кД) – белок пресинаптической мембраны, связанный с кальциевыми каналами; он способен связываться с синаптотагмином. 5. Rab3A (20-25 кД) – белок, препятствующий освобождению медиатора из активной зоны одним квантом. 6. NSF, или N-этил-малеимид чувствительный фактор; это цитозольный белок, который состоит из 4 субъединиц (76 кД); он обладает АТФ-азной активностью и обеспечивает энергией различные внутриклеточные процессы, во время которых происходит слияние мембран и процессы экзо- и эндоцитоза. 7. SNAP – растворимый цитозольный белок, связывающий NSF; он участвует в синаптической передаче после докирования везикулы. 8. SNAP-25 – это белок, связанный с секретосомой. Он расположен на пресинаптической мембране. Вместе с синаптобревином и синтаксином этот белок формирует комплекс, необходимый для экзоцитоза. Все перечисленные белки играют важную роль в процессах экзоцитоза. Все они являются мишенями для ботулинового и столбнячного токсинов.
Функция белкового аппарата, ответственного за экзоцитоз (SNARE-гипотеза). Для объяснения экзоцитоза была предложена SNARE-гипотеза, или гипотеза об универсальной «единице докирования и слияния». Эта гипотеза, как отмечают С.Ю. Черанов и соавт. (1999), предполагает, что в процессе докирования и слияния участвуют такие белки как v-SNARE (или везикулярный SNAP-рецептор), t-SNARE, или мембранный рецептор для SNAP (он является мишенью для v-SNARE), цитозольные белки NSF и SNAP-белок (адаптор к NSF).
Докирование везикулы осуществляется благодаря комплементарности v-SNARE и t-SNARE. Эти два белка-рецептора взаимодействуют друг с другом по принципу «антиген-антитело». После контакта, т.е. когда они свяжутся между собой, оба этих белка становятся рецепторами для SNAP, который в свою очередь присоединяет NSF. При этом комплекс v-SNARE + t-SNARE + Са-канал является осью, на которую нанизываются другие молекулы, участвующие в экзоцитозе. Гидролиз АТФ с участием NSF ведет к пространственной реорганизации SNARE-комплекса, что приводит синаптическую везикулу в состояние прайминга. Вход кальция дестабилизирует SNARE-комплекс везикулы и приводит к экзоцитозу. В целом, последовательность событий от момента прихода везикулы в пул 1 до момента ее прайминга можно представить следующим образом: v-SNARE + t-SNARE → докирование везикулы → v- SNARE + t-SNARE + SNAP → прайминг везикулы.
Роль ионов кальция в экзоцитозе. Нейрональные Са-каналы являются гетерометрическими белками, которые состоят из трех основных субъединиц. В условиях покоя с участием Са-каналов осуществляется докирование синаптической везикулы в активной зоне (это реализуется совместно с белковым аппаратом экзоцитоза, т.е. синтаксином и SNAP-25). За счет такого докирования предотвращаются процессы случайного экзоцитоза синаптических везикул.
При деполяризации пресинаптической мембраны под влиянием потенциала действия Са-каналы открываются, ионы Са2+ входят в пресинаптическое окончание, взаимодействуют с белковым аппаратом экзоцитоза и тем самым инициируют высвобождение медиатора в синаптическую щель. Детальное изучение этого процесса показало, что для осуществления экзоцитоза необходимо создание высокой концентрации ионов Са2+ у везикулы в очень короткий промежуток времени. Короткоживущее «облако» ионов Са2+ около внутреннего устья канала в цитоплазме (с концентрацией более 100 мкмоль/л) получило название кальциевого микродомена. Везикулы могут подвергаться экзоцитозу только в том случае, если их Са-сенсоры находятся внутри кальциевого микродомена. Это достигается за счет связи Са-каналов с докированными синаптическими везикулами. При деполяризации ионы Са через большое количество близко расположенных Са-каналов входят внутрь пресинаптического окончания и создают область повышенной концентрации ионов Са2+ в определенном участке пресинапса, тем самым формируя кальциевый микродомен. Этот домен может включать и несколько соседних, близко расположенных активных зон. В поддержании внутриклеточной концентрации ионов кальция принимают участие различные буферные системы, которые действуют очень эффективно и оставляют только 0,1-2,0 % вошедшего кальция свободным. Подобную функцию выполняют митохондрии и эндоплазматический ретикулюм везикулы. Эти структуры способны не только быстро утилизировать кальций, но и некоторое время поддерживать его высокую локальную внутриклеточную концентрацию за счет освобождения в цитоплазму. Поэтому кальциевый микродомен существует более длительно, чем длится входящий кальциевый ток и обеспечивает более высокий уровень освобождения медиатора в этом месте, что имеет значение в механизмах кратковременной памяти. В целом, ионы Са2+ активируют АТФ-азную активность NSF, благодаря чему совершается контакт везикулы с пресинаптической мембраной и выброс медиатора в синаптическую щель.
Синтез рецепторов постсинаптической мембраны. Рецепторы – это белковые структуры, которые являются интегральными белками плазматической мембраны. Они синтезируются в рибосамах эндоплазматического ретикулюма клетки. После предварительной «сортировки» в аппарате Гольджи они встраиваются в постсинаптическую мембрану. Синтез рецепторов контролируется йодсодержащими гормонами щитовидной железы и, вероятно, другими гормонами, к которым имеются соответствующие гормонсвязывающие сайты на генах, ответственных за синтез данного вида рецепторов. Кроме того, синтез рецепторов определяется наличием медиатора. Например, если мышца денервирована, то число холинорецепторов в области постсинаптической мембраны снижается, но одновременно во внесинаптических областях возрастает концентрация холинорецепторов, что делает мышцу более чувствительной в отношении циркулирующего в крови ацетилхолина. При нарушении синтеза рецепторов или при наличии антител к ним, их концентрация на постсинаптической мембране может существенно снижаться, что уменьшает функциональные возможности синаптической передачи.
Активация постсинаптических рецепторов. Вторичные посредники. Каждый рецептор постсинаптической мембраны имеет сайт (активный центр) для связывания медиатора (агониста). Кроме того, рецептор может иметь сайты для связывания модуляторов или комедиаторов, благодаря чему сродство данного рецептора к медиатору может существенно изменяться. Например, в крови и ликворе есть эндогенный сенсибилизатор β-адренорецепторов (его функцию выполняют гистидин, триптофан, тирозин), повышающий в 10-100 раз чувствительность этих рецепторов к норадреналину и адреналину, а также эндогенный блокатор М-холинорецепторов, снижающий чувствительность М-холинореактивных структур к ацетилхолину.
Для активации большинства рецепторов необходимо более одной молекулы агониста (медиатора). Например, для активации одного холинорецептора нужны две молекулы ацетилхолина, что объясняется наличием двух симметричных агонист-связывающих альфа-субъединиц в составе холинорецептора. Процесс взаимодействия рецептора и медиатора-агониста можно представить в виде схемы: А+Р ↔ АР + А ↔ А2Р ↔А2Р*, где А – агонист, например, ацетилхолин. Р-рецептор в покоящемся состоянии, АР – рецептор, связавший одну молекулу агониста, А2Р – рецептор, связавший две молекулы медиатора, но пока с закрытым состоянием канала, А2Р* – рецептор с открытым (проводящим) ионным каналом.
При активации ионотропных рецепторов, содержащих внутри себя ионные каналы, происходит изменение проницаемости этих каналов, что в конечном итоге приводит к деполяризации или гиперполяризации постсинаптической мембраны. При активации метаботропных рецепторов происходит опосредованное изменение проницаемости ионных каналов, изменение активности ионных насосов или ферментов метаболических процессов в данной клетке. При активации метаботропных рецепторов передача сигнала внутрь клетки осуществляется с участием вторичных посредников – мелких, подвижных молекул (цАМФ, цГМФ, инозитол-3-фосфат, диацилглицерол), способных активировать протеинкиназы (ферменты, переносящие фосфатную группу от АТФ на белок-мишень). Фосфорилированный белок (ионный канал, ионный насос, фермент метаболической цепочки и др.) меняет свою активность и тем самым обеспечивает передачу синаптического сигнала. Образованию вторичных посредников предшествует передача сигнала от мембранного рецептора через ГТФ-связывающий белок (G-белок) к внутриклеточному примембранному ферменту (аденилатциклазе или гаунилатциклазе). При этом G-белок может активировать или ингибировать этот фермент (разновидности этого белка обозначают соответственно как GS-белок и Gi-белок).
Для различных синапсов существует своя последовательность передачи сигнала (медиатор, G-белок, вторичный посредник, протеинкиназа, белок-эффектор). Например, для моноаминов (адреналина, норадреналина, серотонина) она выглядит так: Gs → аденилатциклаза → цАМФ → протеинкиназа А → ионные каналы, ферменты обмена углеводов и др. Для ацетилхолина: Gs → гуанилатциклаза → цГМФ → протеинкиназа Г → калиевые ионные каналы, кальциевые насосы. Для моноаминов и пуринов: Gi → фосфолипаза С → инозитол-3-фосфат/диацилглицерол → протеинкиназа С и рецепторы инозитол-3-фосфата → кальциевый ионный канал. Для пуринов: Gi → фосфолипаза С → арахидоновая кислота → калиевые ионные каналы.
Медиатор. В роли медиатора выступают десятки и даже сотни химических веществ. Для того чтобы назвать вещество медиатором, используют следующие критерии. 1. Вещество выделяется из клетки при ее активации. 2. В клетке имеются ферменты для синтеза данного вещества. 3. В соседних клетках имеются белки-рецепторы, активируемые данным медиатором. 4. Фармакологический (экзогенный) аналог имитирует действие медиатора. Наряду с медиаторами в синапсе есть модуляторы, т.е. вещества, которые прямо не участвуют в процессе передачи сигнала от нейрона к нейрону, но могут, однако, этот процесс существенно усиливать или ослаблять.
Локализация медиаторов и соответствующих нейронов ЦНС. В разных отделах ЦНС расположены нейроны, имеющие различные медиаторы. Ацетилхолин является медиатором альфа-мотонейронов спинного мозга и ствола (их аксоны заканчиваются нервно-мышечным синапсом), нейронов коры больших полушарий, ретикулярной формации мозга, преганглионарных нейронов вегетативной (симпатической и парасимпатической) нервной системы, ганглионарных нейронов парасимпатической нервной системы, а также нейронов сетчатки.
Дофамин является медиатором нейронов, сконцентрированных в среднем мозге (черная субстанция, покрышка мозга), в гипоталамусе, в симпатических ганглиях и в сетчатке. Аксоны дофаминергических нейронов достигают нейронов базальных ганглиев, лимбической системы, коры больших полушарий.
Норадреналин является медиатором нейронов, локализованных в голубом пятне ствола мозга (с проекцией в кору мозга, гипоталамус, мозжечок, спинной мозг), а также ганглионарных нейронов симпатической нервной системы.
Серотонин является медиатором серотониергических нейронов, локализованных, главным образом, в ядрах шва ствола мозга (проекция аксонов в кору головного мозга, гипоталамус, мозжечок, спинной мозг) и в сетчатке.
Гистамин является медиатором нейронов сосцевидных телец гипоталамуса. Аксоны гистаминергических нейронов проецируются в кору мозга, таламус, базальные ганглии, мозжечок, спинной мозг.
Глютамат является медиатором возбуждающих нейронов, локализованных в различных отделах коры больших полушарий, в таламусе, базальных ганглиях, мозжечке, гипоталамусе, стволе мозга, спинном мозге, а также в сетчатке.
Глицин является медиатором тормозных нейронов, расположенных главным образом в спинном мозге и в сетчатке.
ГАМК является медиатором тормозных нейронов коры мозга, мозжечка, ствола мозга, спинного мозга (совместно с глицином) и сетчатки.
АТФ является медиатором возбуждающих нейронов уздечки головного мозга, спинного мозга, афферентных нейронов, а также симпатических нейронов.
Нейропептиды как медиаторы ЦНС. В последние годы в ЦНС выявлено большое число нейроактивных пептидов. Также как и медиаторы (нейротрансмиттеры), пептиды идентифицируются как нейроактивные вещества в соответствии с указанными выше критериями. Для них характерна следующая локализация в ЦНС. Субстанция Р – головной мозг, окончания афферентных нейронов в ноцицептивной системе. Вазопрессин, окситоцин – задний гипофиз, продолговатый мозг, спинной мозг. Кортиколиберин – медиальное возвышение гипоталамуса и другие