На фигурах 1 и 2 схематически показаны деревья дифференсации (Гордон. 1993), которые могли бы описать морфогенез через время у трансформирующихся (метаморфозных) и неотенических саламандр, соответственно. Каждый узел или точка ветвления деревьев представляет собой клеточную дифференцировку. Каждое ребро представляет соответствующий каскад регуляторных генов, структурных и ферментативных белков, а также механизмы контроля на транскрипционном и трансляционном уровнях, плюс их морфогенетические последствия (дифференциальная адгезия, миграция клеток, изменения в морфологии клеток и т.д.).) По сравнению обоих деревьев диффе-ренсации, различия становятся очевидными. Средняя горизонтальная линия соответствует времени нормального метаморфоза. Ветви после метаморфоза сгруппированы в генные каскады: 1) половых клеток и репродуктивных тканей и 2) взрослых соматических характеристик. Так как неотения-это репродуктивная зрелость с отменённым или отложенным соматическим развитием, то "взрослые" ветви для трансформированной особи вообще не экспрессируются у неотенической особи (Фиг.2). Предыдущие ветви просто увековечиваются в течение жизни, как это показано.
Как уже говорилось выше, многие ткани задеваются во время метаморфоза. Поэтому становится очевидным, что, поскольку взрослые гены не экспрессируют у неотеников, они также не подвергаются естественному отбору во время неотении. Тогда мы бы заподозрили, что мутации, происходящие во взрослых генах, не будут удалены естественным отбором до тех пор, пока не произойдет трансформация, и будут накапливаться всё время, пока трансформирующееся состояние не будет восстановлено. Последствия от таких накапливающихся мутаций могут привести к проблемам развития, возникающим при попытках трансформации. Чем больше поколений проходят жизненные циклы без трансформации, тем больше мутаций накапливается. Такие мутации, происходящие во взрослых генах неотеников, будут и дальше способствовать сохранению неотении через положительную обратную связь. Популяции, остающиеся неотеническими в течение длительных периодов времени, могут навсегда запереться в неотеническом состоянии. Этот аспект, возможно, был тем, который рассмотрел Gould (1977), когда он заявил:
"Режим K-селекции благоприятного и относительно постоянного пруда вполне мог бы предложить окончательное подавление неиспользуемой способности метаморфоза."

|
Фиг.2 Дифференсационное дерево неотенических животных. Ни одна из взрослых соматических тканей не появляется, если животное не вынуждено проходить метаморфозу. Если это факультативный неотеник, его дерево дифференциации рассмотрено на Фиг. 1.
В настоящее время нет генетической информации, позволяющей предположить, что эти мутации "взрослых" генов сохраняются в неотенических популяциях. Доказательство с помощью попыток вызвать метаморфоз у неотенических особей инъекцией гормонов гипофиза, однако, обеспечивает некоторую поддержку этой гипотезе. Эксперименты с облигатными неотениками Necturus spp. и Proteus anguinus показали, что их ткани нечувствительны к гормону щитовидной железы (Линн, 1961). Обработка привела дополнительной линьке и небольшому уменьшению жабров (трансформации не произошло). также было заявлено, что увеличение обработки инъекциями тироида была летальна. Мы предполагаем, что клетки Necturus spp. и Proteus spp. чувствительны к тироидному гормону и что активация щитовидной железы принуждает прогрессирование дифференсационного дерева в дефектные взрослые ветви. Как резюмировано выше, эти виды, как полагают, были неотеническими наиболее длительное время среди большинства видов саламандр. Облигатно неотеническая Ambystoma mexicanum легко трансформируется при обработке гормоном щитовидной железы (Taurog, 1974). Также факультативно неотенические Ambystoma tigrinum и Notophthalmus viridescens легко вызвать на трансформацию(Моррис & Герн, 1976; Dent, 1968). Эти виды стали неотеническими гораздо позже, чем Necturus spp. и Proteus anguinus. Летальность у облигатных неотеников может быть обусловлена, следовательно, отключением личиночных генов (и, как следствие, регрессией тканей до апоптоза через лизосомные энзимы и макрофаги), так как дефектные взрослые ветви дифференсационного дерева активируют: генные продукты мутации взрослых ветвей, могущие иметь редуцированную ферментативную активность, дисбаланс ферментных путей и др.. С потерей активности личиночных генов метаболизм организма разрушается. Дисбаланс из-за включения дефектных взрослых генов также может быть смертельным, даже если личиночные ветви продолжают функционировать одновременно. С другой стороны, некоторые взрослые ветви могут быть индуцированы без запуска других соседних ветвей, производя метаболическую несовместимость.
Дифференсационные деревья могут также объяснить, почему облигатные неотениик имеют так много ДНК, доселе неразгаданную тайну (Duellman and Trueb, 1986). Для ветвей дифференсационного дерева, которые используются, генетическая дупликация ветви может быть вредна вследствие эффектов генетической дозировки (Дибан и Баранов, 1987), если баланс не достигается за счет общей полиплоидии генома, отличительной черты саламандр (Fankhauser, 1955; Lowcock and Murphy, 1991). Для неиспользованных взрослых ветвей дифференсационного дерева нет явных ограничений на дупликацию, кроме изменения в физиологии, которое происходит с увеличением размера клеток. Так размер ДНК может увеличиваться неконтролируемым образом до тех пор, пока не будут достигнуты пределы, лимитирующие клеточный размер (тем не менее, cf. Larson, 1984). Давно известно, что размер генома может влиять на морфологический и физиологический фенотип клеток (Кавалье-Смит, 1978, 1985). Olmo и Morescalchi (1975, 1978) показали, что у хвостатых и бесхвостых амфибий существует прямая положительная корреляция между размером генома и размером клетки. Было предположено, что больший размер клеток коррелирует с более низким метаболизмом (Szarski. 1970; Duellman and Trueb, 1986) и следовательно, предполагает более длительное развитие, которое наиболее возможно при более низких температурах. Goin et al. (1968), Хорнер и Макгрегор (1983), Sessions и Larson (1987) и Пейджел и Джонстон (1992) скоррелировали большой размер ядерного генома с более медленными темпами развития. Кроме того, в условиях холодной воды факультативные неотеники наблюдались Sprules (1974a, b), наводя на размышления, что по крайней мере, стартовые условия обитания как облигатных, так и факультативых неотеников одинаковы. (Облигатные неотеники столкнувшись с изменениями условий окружающей среды, должны либо приспособиться к изменившейся водной среде либо вымереть, в то время как факультативные неотеники имеют возможность трансформации. Таким образом, облигатные неотеники должны быть обнаружены в несколько большем диапазоне водных сред, чем неотенические формы факультативных неотеников). Licht и Lowcock (1991) напоминают нам, что мы должны быть осторожны, когда пытаемся провести корреляцию между размером генома и метаболизмом всего организма. Хотя хвостатые амфибии имеют самый низкий уровень метаболизма всего организма из всех тетрапод, в их обширном анализе, Licht и Lowcock (1991) не смогли продемонстрировать никакой корреляции между нормами метаболизма и размером генома за исключением неотенических видов Necturus и Amphiuma. Интересно, что они считаются двумя из самых старейших неотенических родов хвостатых амфибий. Как они заявляют, влияния увеличения размера генома увеличивающие результаты недавней полиплоидизации, не обязательно будут такие же, как произошедшие в геноме дупликации (как мы предположили для неотеников; cf. Cavalier-Smith, 1985) и что адаптация к геномным, нуклеотипическим и организменным эффектам будет ожидаться только после очень длительного давления отбора.
Может ли этот сценарий быть коррелирован с другими основными дупликациями генов неизвестной функцией (мусорная ДНК: Ohno, 1972; John and Miklos, 1988), еще предстоит выяснить. Дублирование неиспользуемых взрослых генов у облигатных неотеников представляет собой альтернативу провалившейся гипотезе "множественных амплификаций с последующей дисперсией продуктов амплификации по всей длине всех хромосом" (John and Miklos, 1988). Наша гипотеза предполагает, что мРНК, уникальные для взрослой ткани трансформированных саламандр, должны иметь больше мест, связующих с ДНК, для геномов облигатных неотеников с гаплоидным геномом, чем для остальных саламандр. Единственным фактором, работающим против этого, был бы распад дулицированной ДНК на псевдогены через мутацию (Li and Graur, 1991) до такой степени, что комплементарная привязка была бы утрачена. Поэтому более поздние облигатные неотеники лучше подходят для этого эксперимента. С другой стороны, у них может быть меньше дупликаций во взрослых ветвях дифференсационного дерева. Подобный эксперимент был проведен Morescalchi и Serra (1974) путём определения кривизны изгиба у ДНК неотенических и обычных хвостатых амфибий на основе того факта, что наибольшая кривизна у наиболее часто повторяющихся последовательностей ДНК. Компоненты часто повторяющихся последовательностей наиболее встречаемы у неотенических видов. Эти повторяющиеся последовательности могут дублироваться неиспользуемыми взрослыми ветвями. Это также предполагает возможное родство между длиной существования облигатного неотенического вида и размером генома (Фиг. 3).
Датирование ядерной ДНК посредством геологической продолжительности неотении, использующей таксоны семейств, была собрана Duellman и Trueb (1986) и подробной компиляции, Licht и Lowcock (1991). Amphiumidae и Sirenidae, по этим данным, самые древние неотенические хвостатые с их наиболее вероятным возникновением в верхнем мелу (135 m.y.a). Cryptobranchidae, Dicamptodontidae и Proteidae известны из палеоцена (65 м. y.a). Семейство Salamandridae известно из эоцена (50 м. y.a). Неотенические хвостатые амфибии с самым маленьким размером генома, Ambystomatidae и Plethodontidae, сформировались совсем недавно, в течение Олигоцена (35 m.y.a) и Миоцена (23 m.y.a), соответственно. Корреляция может достаточно хорошо вписываться в прямую линию (ДНК [pg] = (0.6299 pg/1000000 лет) возраст [1000000 лет] + 53.4 pg) (r=0.5298) (Фиг. 3). Если экстраполировать к нулевому геологическое время, то получим примерное количество ДНК, найденной в нон-облигатных неотенических саламандрах: 53.4 pg / ядро (это Вычисленное количество!). Средняя ядерная ДНК, наблюдаемая (а это Наблюдаемое количество!) в необлигатных саламандрах, составляет 55 плюс-минус 20 pg / ядро (Таблица 16.2 у Duellman и Trueb, 1986), что является замечательным согласием. Эта корреляция подтверждает палеонтологическую летопись и морфологический филогенетический анализ у Duellman и Trueb (1986).
DlSA/Nucleus vs. Evolutionary Duration for Families of
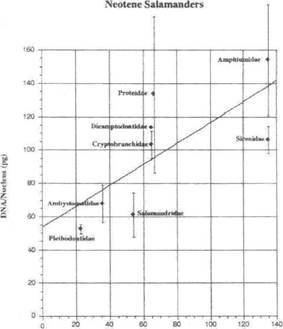
Фиг.3 Размер генома по сравнению с эволюционной продолжительностью для неотенических семейств саламандр. Прямая линия экстраполяции показывает, что, как представляется, существует возможная корреляция между тем, как долго вид саламандр был облигатно неотеническим и сколько ДНК он накопил. Это говорит о возможности появления вторых молекулярных часов, включающих скорость дупликации мусорной ДНК (погрешность такта =стандартное отклонение).
Некоторые последние молекулярные данные указывают на более раннюю радиацию большинства видов, чем указывалось в геологической летописи (ср. Ларсон и Диммик. 1993). Однако, поскольку только филогенетически примитивные семейства, по-видимому, имеют наибольшее количество ядерной ДНК, отсутствие более ранних окаменелостей послужило бы сдвигу в Фиг. 3 к более крутой линии.
Дублирование ДНК не должны быть более крупных порций взрослых дифференциальных ветвей (см (cf). все удвоения генома (Sparrow и Науман, 1976)), но, это, возможно, непрерывный процесс дупликации индивидуальных ветвей. Если увеличение ДНК у облигатных неотениках действительно дублирует "взрослую" ДНК, то сравнение ДНК факультативных и облигатных неотеников может дать карту размещения взрослых ветвей дифференсационного дерева на хромосоме.
Облигатные неотеники, таким образом, наглядно демонстрируют экофизиологическую роль мусорной ДНК: селекция по норме низкого метаболизма может возникать через селекцию на более крупный клеточный размер, что требует больше ДНК. Предположение о связи между размером генома, размером клетки и метаболизмом было предложено раньше (Szarski, 1983), подключение палеонтологических доказательств (Фиг. 3) может быть новостью.
Способность allophenic-комбинаций нормальных и анеуплоидных млекопитающих для "спасения" анеуплоидных клеток или даже летальных мутаций, с точки зрения нормального развития (Айкер и Хопп, 1973; Макларен, 1976; Dyban и Баранов, 1987) свидетельствует о возможности похожих экспериментов на allophenic-комбинации облигатных и факультативно неотенических саламандр. Можно предположить, что результирующие особи будут факультативно неотеническими, по аналогии с экспериментами на млекопитающих.
Очевидно, что существует широкий спектр использования неотении. Облигатные неотеники никогда не трансформируются в естественных условиях, а вынужденная трансформация оказывается летальной, тогда как факультативные неотеники могут либо трансформироваться, либо оставаться в личиночном состоянии. Неотенические и трансформированные особи факультативных неотеников могут встречаться даже в одних и тех же популяциях. Наблюдается также диапазон индуктивности трансформации у саламандр, в зависимости от длительности выраженного неотенического состояния. Возможно, за многие годы облигатно неотенические Ambystoma mexicanum могут стать неиндуцибельными. К сожалению, считается, что их исчезновение неизбежно (Armstrong and Malacinski 1989). В любом случае, мы полагаем, что путь превращения от: факультативной неотении => к индуцибельной облигатной неотении и => к неиндуцибельной облигатной неотении представляет результат работы ДВУХ молекулярных часов:
1) известные молекулярные часы работают в точках замещения (Li и Grauer, 1991);
2) молекулярные часы, определяются скоростью дупликации ветвей взрослой части дифференцирующего дерева.
Последние часы могут перестать работать после некоторого геологического времени, если есть экофизиологические ограничения на общее количество ДНК в отдельно взятой клетке, хотя Фиг. 3 показывает, что этот предел еще не достигнут.
Вышеуказанные стадии в прогрессировании неотенического фенотипа можно охарактеризовать следующим образом:
1) факультативная неотения: действуют только факторы окружающей среды, такие как условия температуры, света и воды, изменяя гормональную активность и вызывая метаморфоз или неотению. В этих условиях все ткани годятся для метаморфоза и успешного дифференцирования в взрослые ветви дифференсационного дерева. Примеры: Notophthalmus vridescens и Triturus cristatus.
2) индуцибельная облигатная неотения: экологические и генетические факторы действуют вместе, чтобы определить прогрессирование тканевой дифференсации. Состояние облигатной неотении поддерживается в природе снижением чувствительности (через генетические изменения) к эффекторам метаморфоза (экологические и гормональные эффекторы). Опять же, все ткани ещё могут быть преобразованы, но пороговые уровни для начала действия сигнала дифференсации увеличиваются. Пример: Ambystoma mexicanum.
3) неиндуцибельная неотения: вмешиваются только генетические факторы, предотвращая тканевую дифференцировку и метаморфоз. Взрослые ветви дифференсационного дерева изменяются до такой степени, что дифференсация в эти взрослые ветви становится неблагоприятной или летальной. Поскольку дифференсация в эти взрослые ветви неблагоприятна, селекция может вести к тому, что узлы дифференсационного дерева становятся нечувствительными (при нормальной стимуляции) к сигналу метаморфоза. Ткани организма в этом случае могут быть не вполне комплектными. Это условие может рассматриваться не как истинная неотения, поскольку по определению она подразумевает возможность для не-личиночной репродуктивной стадии. Неиндуцибельные или облигатные неотеники конечно не обладают такой жизненной стадией. Примеры: Necturus и Proteus.
Все вышеприведенные утверждения поддерживают идею о том, что неотения может играть значительную роль в качестве механизма изолирования популяции и далее в качестве механизма видообразования у саламандр. Данные будут поддерживать менее специфические представлении об организации генома, чем те, которые укладываются в концепт дифференсационных деревьев. Несмотря на это, помимо возможности того, что они являются точным представлением хода развития, мы находим дифференсационные деревья отличной моделью для генерирования идей о связях между развитием, генетикой, эволюцией и экологией. Единственная другая модель общей организации генома является неиерархической (Kauffman, 1971, 1973, 1993).
Резюме
Как было показано ранее, неотения описывается как достижение личиночного размножения через отложенное соматическое развитие. Эксперименты предполагают, что неотения генетически наследуема, и такие генетические вариации неотении присутствует в большинстве популяций (в меньшей степени у облигатных неотеников). Эти эксперименты также показали, что факторы окружающей среды являются мощным селективным прессом для регулирования частоты случаев неотении и что быстрая фиксация неотенического характера может быть вызвана у последующих поколений в относительно короткий период времени. Мы предположили, что не существует селективных сил, которые могли бы воздействовать на взрослые гены неотенических популяций, что приводит к накоплению мутаций во "взрослых" генах. Доказательства "принуждения к неотении", кажется, подтверждают эту гипотезу.
В последовательности "факультативные неотеники=>вынужденно облигатные неотеники => невынужденно облигатные неотеники" размер клетки и содержание ДНК в клетке увеличивается, несомненно, сопровождается снижением метаболической активности. Мы предполагаем, что это показывает функцию для дополнительной, нефункциональной, так называемой "мусорной" ДНК. Это также предполагает работу вторых молекулярных часов, связанных со скоростью дублирования неиспользуемой ДНК, которая может быть дуплицированна в неиспользуемых взрослых ветвях дифференсационного дерева. Бремерманн (1987) выдвинул альтернативную гипотезу о том, что:
"...Некодирующая ДНК может обеспечить места вставки для ретровирусов, которые будут предотвращать их дальнейшую транскрипцию и репликацию и, таким образом, их распространение будет абсорбировано, примерно так же, как стержни поглотителя впитывают нейтроны в ядерном реакторе."
Он не предполагал, что это приведет к вторым молекулярным часам, хотя это может быть связано.
Молекулярные часы мусорного ДНК свободно работают при трёх предположениях:
1) Все части ДНК подвержены периодической дупликации (сf. Ohno, 1970).
2) Дуплицированная ДНК, которая не используется, также не селектируется, за исключением того, что она изменяет метаболизм, изменяя размер ядра и размер взрослых клеток.
3) Экологические условия, благприятные для более низкого метаболизма, связаны с более крупными клеточными размерами создают селективное давление на мусорную ДНК.
Когда молекулярные часы мусорной ДНК идут свободно, ДНК прибавляет по норме приблизительно 63 pg на 100 миллионов лет (Фиг. 3). Поскольку мы еще не знаем, какая пропорция генома саламандры находится во взрослых ветвях дифференсационного дерева, мы пока не можем нарисовать цифру того, как часто неиспользуемая ДНК дублируется. Однако, подсчитав видовое мРНК у факультативных неотеников на их трёх основных стадиях, можно обеспечить необходимую оценку и, таким образом, калибровку часов.
При использовании дуплицированной ДНК, как в личиночных, так и в репродуктивных тканях у всех саламандр (Фиг. 1 и 2), могут произойти следующие события:
1) Благоприятные мутации при копировании, возможно, дающие новые функции.
2) Нейтральные мутации, добавляющиеся в мусорную ДНК.
3) Неблагоприятные мутации, нарушающие функционирование генов в оригинальной копии.
В третьем случае существует селективный недостаток дупликации ДНК. В простом случае ферменты или белки цитоскелета, которые взаимодействуют на несколько субъединиц необходимы для функционирования. Мы предполагаем, что этот механизм, в дополнение к метаболическому принуждению, из-за клеточного размера, может быть тем, что лимитирует количество мусорной ДНК, образованной из использованной ДНК. Облигатно неотенические саламандры поэтому представляют собой необычный случай, когда большая часть ДНК, а именно взрослые ветви дифференсационного дерева, внезапно переходит в неиспользованное состояние, для которого этот механизм не применяется.
Теперь о критическом (важнейшем) компоненте, отсутствующим в понимании эволюционной значимости неотении у саламандр. Это то, какова же природа генов, контролирующих неотению, и какие именно генетические различия существуют между облигатными и факультативными неотениками. Пока на эти вопросы отвечают исследования неотении, имеющие значительные ограничения и носящие, в основном, описательный характер. Концепция дифференсационного дерева может не дать единственного объяснения, но она дает нам работающую модель иерархической дифференциации тканей организма, в рамках которой такие вопросы могут быть заданы.
Ссылки на литературу
Андерсон, J. P. и Д. р. Worthmgton (1911), "История жизни мексиканской саламандры Ambystoma ordinarium". Тейлор. Герпетология 27, 165-176.
Андерсон, С. л. Н. 3 ами Дент (я 982). Повышенная потеря тироксина из кровотоков тритонов после введения пролактина баранины. Ген. Комп. Эндокринол. 47, 276 274.
Армстронг, Дж, В, И г. М. Mataciiiski (ЭЦП.). (1989). Биология развития, аксолотлей. Издательство Оксфордского Университета, Нью-Йорк.
Bonner, J, T. (1965). Молекулярная Биология развития. Oxford University Press, Oxford.
Бремерманн, Н. И. (1981). Адаптивное значение сексуальности. Expertentia 55(Внешн): 135 161.
Cavalier-Smith, T. (ed.) (1985). Эволюция размера генома. John Wiley and Sons Ltd., Нью Йорк.
Дент. Дж. Н. (1968). Обзор метаморфоза амфибий, с. 271 -311.
Derby, A - (1975). Влияние пролактина и тироксина на резорбцию хвоста Rana pipiens: in vivo и in vitro. Ж. Эксп. / / Зоол. 193: - в 20.
Duellman, е. В. и л. Trueb. (1986). Биология земноводных. McGraw-Hill Book Co, New York.
Данн, Э. Р. 11931). Текущее состояние проблемы аксолотля. Анат. Rec, 51 (Suppl.): 90.
Dyban. А. П. и в. С. Баранова. (1987). Цитогенетика эмбрионального развития млекопитающих, Clarendon Press, Oxford.
Eicher E. М., Р С. Hopp. (1973), Использование химер для передачи летальных генов мыши и демонстрации аттелизма у двух Х-хромосомных самцов летальные гены jp и msd..). Опыт. / / Зоол. 183: IK I -184.
Etkin У. и А. г. гона. (1967), Антагонизм между пролактином и тироидным гормоном в развитии амфибии. 3. Exp. / / Зоол. 165: 249-258.
Figel С. R. и Р, Д. Semlitcsh. (1990). Выживаемость популяционных вариаций и метаморфозы личиночных саламандр (Ambystoma maculatum) в присутствии хищных рыб, Copeia 1990 (3): 818 826
Garstang, W, (1966). Аксолотль и аммокует. p. 62. В Личиночных формах и других Зоологических стихах. Василий Блэкуэлл, Оксфорд, Соединенное Королевство.
Goin,?. В., Осип. С. Ж. и К, Бахман. (1968), ДНК в истории жизни земноводных, Copeia 1968(3): 532-540.
Харрис. Р. Н. (1987). Зависимость педоморфоза от плотности посадки у саламандр Notophthalmus viridescens dorsalis. Экология 68: 70S -712.
Хили, Р. У. (1970). Сокращение неотении в Массачусетской популяции Notophthalmus viridescens. Copeia 1970: 578-581
Хордер, Т. Я. (1983). Эмбриологические основы эволюции, с. 315-352. В... С. Гудвин, холдер Н., И С. C, Wylie, (eds,) " развитие и эволюция. Cambridge University Press, Cambridge.
Jacobs, г. Ф. М., р. С. А. Michielson и Е, р. Kuhn, (1988). Тироксин и трийодотиронин в плазме и щитовидной железе у неотеников и метаморфизованных аксолотлей Ambystoma mexicanum: влияние ТРГ инъекций, быт типа. Эндокринол 70: 145-151,
Кауфман, С. А (1971), сети регуляции генов: теория их глобальной структуры и поведения. Актуальные темы в биологии развития 6: 145-182.
Кауфман, С. А. (1993). Происхождение порядка: самоорганизация и селекция в эволюции. Издательство Оксфордского Университета, Нью-Йорк,
Larson, A. (1984). Neontelogical вывод эволюционных паттернов и процессов у саламандр семейства Plethodontidae. Ф. т. БИОЛЬ. 17: 119-217.
Lamm, A. and W W. Dimmick (1993). Филогенетические отношения семейств саламандр: анализ конгруэнтности морфологических и молекулярных признаков. Herpeto!. Моногр. 7: 77-93,
Ли. У. и Д. Граур (1991). Основы молекулярной эволюции, Sinauer Associates, Сандерленд. Массачусетс.
Лихт, л. Е. и л. А. Lowcock. (1991). Размер генома и скорость метаболизма в саламандрах. Типа. Биохимия. Физиол, 100Щ1): 83-92,
Lowcock, Л, И р. У. Мерфи. (1991). Пентаплоидия в гибридных саламандрах демонстрирует повышенную толерантность к многохромосомным наборам, Experientia 47: 490-497.
Lynn, У. г. (1961), Типы метаморфоз амфибий. Амеры. / / Зоол. 1: 151-161.
Мартин,... C, и R. Gordon. (1993). Ультра-Структуральный анализ состояния клеточного сплиттера в дифференцирующей эктодерме клеток у эмбрионов аксолотля Ambystoma mexicamum, как он представлен.
McLaren, A. (1976). Химеры млекопитающих, Cambridge University Press. Кембридж.
Morescalchi, А. и "в. Серра. (1974). Кинетика ренатурации ДНК у некоторых педогенетических urodelus. Experientia 30: 487-489.
Morescaiehi, А. (1975). Эволюция хромосом у хвостатых земноводных. Эвол. БИОЛЬ. 8: 339 - 387.
Нуссбаум, А. Р. (1976). Географическая изменчивость и систематика саламандр рода Dicamptodon Strauch (Ambystomatidae). Смешанная, Опубл. Mus. / / Зоол. Univ, Мичиган 149: 1-94,
Ohno, S. (1970). Эволюция путем Дупликации генов. Издательство Springer-Veriag, Нью-Йорк.
Ohno, S. (1972). Как много "мусорной" ДНК в нашем геноме, с. 366-370. Гп х. х. Смит, (изд.), Evolution of Genetic Systems, Gordon and Breach, New York.
Olmo, е. А. Morescalchi. (1975). Эволюция генома и размеров клеток у саламандр, Expcrientia 31: 804-806.
Olmo, е. А. Morescalchi. (1978). Геном и размер клеток у лягушек: сравнение с саламандрами. Expcrientia 34: 44-46.
Остер, г. и П Alberch. (1982). Эволюцию и бифуркации в программах развития вузов. Эволюция 36(3):
Пагель, М. и р. А. Джонстон. (1992). Вариации между видами по размеру ядерного генома поддерживают объяснение парадокса с-значения в спам-ДНК. Процесс. R Sot. Б. Бонд 249 119-124
Паттерсон, С. (1983). Чем документы фитогенеза отличаются от онтогенеза? стр. I 31. В... С. Гудвин. N, держатель и. С. Wylic (eds.j, развитие и эволюция. Cambridge University Press. Кембридж.
Пирс, Б. А. и Х. М. Смит. (1979). Неотения или педогенез? 1. Herpetol. 13; 119-121.
Рафф. Р. А. и Т. С. Кауфман. (1983). Эмбрионы, гены и эволюция. Эволюционно-генетическая основа эволюционных изменений. Maemiltan, New York.
Семлич. Д. р. и У. Дж. Гиббонс. (1985). Фенотипическая изменчивость у метаморфозных и педоморфозных саламандр Ambystoma talpoideum. Экология 66: 1123-1130.
Semliwch, р. Д. и Х. М. Уилбур. (1989). Искусственный отбор для pпедоморфозных саламандр Ambystoma talpoideum. эволюции 43(1): Ш5-И2.
Sessions S. K. и A, Ларсон (1987), Корреляты развития размера генома у безлёгочных саламандр и их последствия для эволюции генома, Evolution 41' 1239 1251.
Снайдер, р. С. (1956), Сравнительные особенности истории жизни у Ambystoma gracile (Baird) из популяций на низких и больших высотах. Copeia 1956(1): 41-50.
Плевков, в. г, (1974a). Адаптивное значение педогенеза у североамериканских видов амбистом (амфибии; Каудаты): гипотезы. Мочь. Ж. / / Зоол. 52: 393
Sprules, W.G. (1974b). Факторы окружающей среды и встречаемость неотении у Ambystoma gracile (Baird) (амфибий; Хвостатые). Мочь. Ж. / / Зоол. 52: 1545-1552.
Стюард. Джей Ш. (1969). Хвостатые земноводные Европы, с. 145 148. David and Charles < Publishers) Limited, Великобритания.
Szarski, Х. (1970). Изменение количества ДНК в ядрах клеток в процессе эволюции позвоночных. Природа 226: 651 652.
Szarski, H (1983). Размер клеток и концепция расточительных и экономных эволюционных стратегий. J theor. Biol 105: 201-209.
Tata, J. R (1984). Действие гормонов роста и развития: эволюционные аспекты пп, 1 -58. в R F. Голдбергер и К. р. Ямамото (eds,). Биологическая регуляция и развитие Vol. 3B. Plenum, New York.
Тата, Ж. Р, Кавахара, A и Б. С. Бейкер. (1991). Пролактин блокирует зарождение вызванного морфогенезом тироидного гормона и вызывает клеточную смерть в культурах тканей личинок амфибий. Разработка. БИОЛЬ. 146: 72 80.
Томсон, С. К. (1972). Предпринята попытка реконструировать эволюционные изменения содержания клеточной ДНК двоякодышащих рыб. Ж. Эксп / / Зоол. 180: 363 372.
Томпкинс. Р. (1978), Генетический контроль (Джин-контроль))))метаморфоза аксолотля Amer. / / Зоол. 18: 313-319