Биологическая роль неорганических веществ (О2, СО2, N2 и т. д.), а также ионов металлов и соединений, включающих эти ионы и атомы металлов, связанные ковалентно, чрезвычайно важна и многообразна. Один из основных процессов, связанных с участием неорганических соединений, - процесс фотосинтеза. Именно взаимодействие Н2О и СО2 - двух неорганических соединений - приводит (при катализирующем действии хлорофилла - магниевого комплекса порфирина) к синтезу крахмала С6Н12О6 с выделением кислорода:
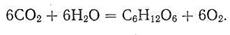
Эта реакция проходит через большое число стадий, но суть фотосинтеза состоит все же в соединении неорганических веществ в углевод. Углеводы, синтезируемые в растениях, потребляются травоядными животными. В результате пищеварения и дальнейших сложных превращений в организме травоядных образуются белки и жиры, служащие пищей для хищников и всеядных животных. К числу последних может быть отнесен и человек. Источником энергии, необходимой для жизнедеятельности человека и животных, являются белки, жиры и углеводы, содержащиеся в продуктах питания растительного и животного происхождения. Окисляясь (неорганическим) молекулярным кислородом, эти вещества, вернее продукты их деструкции, дают организму энергию, затрачиваемую на другие жизненно важные химические процессы и превращаемую в механическую энергию (движение), электрическую энергию и др.
Уже упоминалось, что важнейшую роль в процессах метаболизма играют ионофоры, регулирующие содержание акваионов щелочных и щелочноземельных металлов во внеклеточном и внутриклеточном пространстве, а также ферменты. Ферменты - всегда белковые вещества. Они ускоряют или ингибируют важнейшие процессы в организме, способствуют переносу в тканях кислорода и углекислого газа, переносу электронов, ускорению гидролитических процессов.
Роль акваионов металлов и металлсодержащих комплексов в живых организмах состоит в регуляции процессов, связанных с получением, преобразованием и распределением в организме энергии, выделением вредных для организма продуктов реакции.
Рассмотрим на ряде примеров биологическую роль акваионов металлов и их комплексов с биолигандами.
Транспорт ионов металлов и других неорганических компонентов в растительных и животных организмах
Для нормального функционирования живых организмов необходимо строго определенное распределение химических веществ по различным частям организма. В тканях млекопитающих действует система транспорта ионов натрия или калия, получившая название «натриевый (или калиевый) насос». Важнейшую роль играет также транспорт железа и других биометаллов.
Натриевый насос обеспечивает необходимое соотношение концентраций ионов Na+ и К+ во внеклеточном и внутриклеточном пространстве. Установлено, что в большинстве клеток животного организма [К+] составляет 0,12-0,16 моль/л, тогда как [Na+] в тех же клетках не превышает 0,01 моль/л. Во внеклеточной жидкости соотношение обратное: [Na+]≈0,15 моль/л, а [К+]меньше, чем 0,004 моль/л. Таким образом, существует значительный градиент концентраций ионов К+ и Na+ между внеклеточным и внутриклеточным пространством. Такое распределение не может быть самопроизвольным, очевидно, требуется затрата энергии, чтобы ионы К+ накапливались внутри клеток, а ионы Na+ оттуда выводились. Установлено, что «насос», накачивающий ионы К+ в клетки к выкачивающий оттуда ионы Na+, т. е. действующий против концентрационного градиента, работает с помощью фосфатопротеина, который образует с ионами К+ более прочные соединения (в силу соответствия размеров иона К+ и полости ионофора), чем с ионами Na+. В составе комплекса с фосфатопротеином ион К+ проходит через клеточную мембрану. Во внутриклеточном пространстве фосфато-протеин взаимодействует с аденозинтрифосфатом. Новый лиганд образует более прочное соединение с ионами Na+, нежели с ионами К+ и выводит ионы Na+ из клетки во внеклеточное пространство.
Железо в виде ионов Fe2+, Fe3+ и биокомплексов необходимо организму животных и человека для выполнения важнейших жизненных функций, таких, например, как перенос кислорода и катализ окислительно-восстановительных процессов, служащих одним из главных источников энергии. В организме животных и человека железо накапливается и сохраняется в печени, селезенке и костном мозге в виде белковых образований - ферритина и гемосидерина. Белковая часть ферритина представляет собой сферическое образование (внешний диаметр около 120 Ǻ) с внутренней полостью диаметром около 75 Ǻ. В полости ферритина находится мицелла, состоящая из гидратированного и гидролизованного фосфата Fe(III). Масса сухого остатка, содержащего железо, составляет 23% от массы ферритина (в гемосидерине железа еще больше).
Транспорт железа от ферритина к красным кровяным тельцам происходит с помощью белка трансферрина, очень прочно связывающего Fe (III) в хелат. Установлено, что хелаты Fe(III), образованные белками типа трансферрина, переносят железо через биомембраны, тогда как сам по себе фосфат железа, содержащийся в ферритине и гемосидерине, преодолеть этих преград не может.
Транспорт О2 в организме животных и человека осуществляется железосодержащими комплексами - гемоглобином и миоглобином. Оба этих белка содержат «гем-группы», представляющие собой порфириновый комплекс железа:
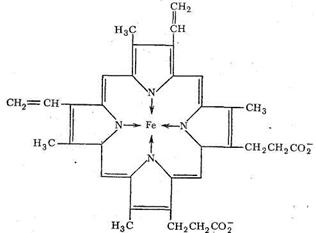
Гемовая группа присоединяется к белковой части молекулы гемоглобина и миоглобина путем координации гистидинового атома азота белка ионом железа гемовой группы. Таким образом, в координационную сферу иона железа входит пять атомов азота (четыре атома азота порфирина, один атом азота белкового гистидина). Шестое координационное место занимает Н2О или О2.
Гемоглобин имеет молекулярную массу 64 500 и включает четыре гем-группы. Миоглобин сходен с гемоглобином по строению, но содержит только одну гем-группу. Гемоглобин почти так же хорошо, как миоглобин, связывает кислород при высоком давлении кислорода, но, когда давление кислорода падает, преимущество в связывании О2 имеет миоглобин. Падение давления О2 наблюдается в тканях мышц, потребляющих кислород. Результатом использования кислорода является накопление в мышцах СО2 и, как следствие, понижение рН. Это еще более способствует высвобождению О2 из гемоглобина и передаче кислорода миоглобину. Таким образом, и гемоглобин, и миоглобин участвуют в переносе кислорода.
Теряя кислород, железо в гемоглобине (форма дезоксигемоглобина, КЧ железа = 5) переходит в высокоспиновое состояние. Ослабление связи с атомами азота порфирина приводит к выходу атома железа из плоскости кольца порфирина (на 0,7-0,8Ǻ). Напротив, вследствие присоединения кислорода к гемоглобину образуется низкоспиновый октаэдрический комплекс, железо в котором находится в плоскости порфиринового кольца (КЧ железа = 6). Хотя связывание молекулярного кислорода гемоглобином и миоглобином сопровождается ослаблением связи O = O в молекулярном кислороде из-за размещения π-дативных электронов железа на разрыхляющих орбиталях О2, этот процесс сам по себе не сопряжен с необратимым переносом электрона, поэтому его обычно не причисляют к ферментативным окислительно-восстановительным реакциям. Отметим, однако, что разрыхление связи O = O в не может не активировать окислительные реакции.
Например, можно рассматривать взаимодействие активированной формы О2 с восстановленной формой субстрата StH2, сопровождающееся переходом его в окисленную (дегидрированную) форму St:
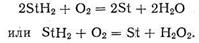
|
При этом кислород превращается в Н2О и Н2О2 с выделением энергии.
Металлсодержащие ферменты
Далеко не все ферменты, катализирующие биологически важные процессы в организме животных и растений, содержат ионы металлов. Однако металлсодержащих ферментов только в организме человека несколько сотен, и их биологическая роль чрезвычайно важна. Нарушение структуры или удаление из организма даже одного из них приводит к тяжелым расстройствам функций организма, а затем и к его гибели.
Металлсодержащие ферменты представляют собой координационные соединения, и поэтому их исследованием занимается неорганическая химия или, точнее, координационная химия.
Рассмотрим строение и биологическую роль нескольких металлоферментов.
Каждый фермент состоит из двух частей: кофермента и апофермента. Кофермент - это легко отделяемая часть фермента. В металлоферментах ион металла составляет основную часть кофермента. Апофермент-остальная часть фермента, которая в отсутствие кофермента не проявляет ферментативной активности.
Обычно рассматривают две группы металлоферментов, различающихся по типу катализируемых ими реакций: ферменты, катализирующие реакции гидролиза, и ферменты, катализирующие протекание окислительно-восстановительных реакций.
Ферментативный катализ реакций гидролиза
Реакции, протекающие в организмах животных и растений с участием воды, играют важнейшую роль в процессе обмена веществ и служат одним из основных источников "энергии, запасаемой организмом.
Реакции гидролиза катализируются обычно теми металлоферментами, которые содержат ионы Са2+, Mg2+, Zn2+ и Мn2+. Хотя ион Мn2+ легко вступает в окислительно-восстановительные реакции, в ходе гидролитических реакций он, как и все упомянутые ионы, не меняет степени окисления.
Наиболее хорошо изучены строение и биологическая роль двух из металлоферментов, катализирующих гидролитические процессы, - цинксодержащих ферментов карбоангидразы и карбоксилазы. Отметим, однако, что только в организме человека содержится несколько десятков цинксодержащих ферментов, различающихся по своему строению и функциям.
Карбоангидраза состоит из больших белковых молекул с молекулярной массой 430 000. Каждая из таких макромолекул содержит 260 аминокислотных остатков. Форма молекулы карбоангидразы - эллиптическая, размеры ее (40x45x55) А3. Эллиптическая молекула карбоангидразы имеет полость, причем аминокислоты, составляющие белок апофермента, расположены так, что гидрофобные их части как бы выстилают внутреннюю полость макромолекулы. В одной молекуле фермента содержится только один ион Zn2+, который находится вблизи центра полости карбоангидразы. Координационный полиэдр комплекса, центральным ионом которого является Zn2+, представляет собой искаженный тетраэдр. Три положения в координационной сфере Zn2+ занимают донорные атомы азота из имидазольных групп аминокислоты гистидина:
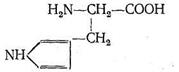
Четвертое координационное место занято молекулой Н2О или ионом ОН-. Фермент карбоангидраза содержится в красных кровяных тельцах. Установлено, что карбоангидраза примерно в 106 раз ускоряет реакцию образования бикарбонат-иона и обратную ей реакцию распада НСО3-:
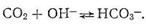
Рассмотрим предполагаемый «концертный» механизм действия карбоангидразы. Ион Zn2+, входя в состав несимметрично построенного координационного соединения (имеющего по этой причине напряженную структуру), обладает высокой реакционной способностью. Столкновение на активном центре карбоангидразы иона Zn2+, СО2 и иона ОН - в полости карбоангидразы приводит к активации всех участвующих в реакции веществ, в результате чего быстро образуется бикарбонат цинка:
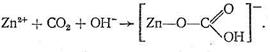
Следующий этап процесса состоит в обмене образовавшегося бикарбонат-иона на Н2О или ОН-, т. е. в высвобождении активного центра фермента для последующих актов катализа.
Установлено, что ингибирование реакции синтеза и распада бикарбонат-ионов достигается введением в координационную сферу иона Zn2+ лигандов, образующих более прочную связь с Zn2+, чем с НСО3-,Н2О или ОН-. Такими являются ионы CN-, N3-, S2O42 - и др. Так как в некатализируемых условиях (в присутствии CN-, N3 - и т. д.) синтез НСО3 - и его распад протекают слишком медленно, физиологические требования не выполняются, нормальное состояние организма нарушается.
Каталитическая активность карбоангидразы не понижается при замене Zn2+ на ион Со2+, в других случаях активность фермента либо падает, либо исчезает.
Карбоксипептидаза представляет собой фермент, вырабатываемый поджелудочной железой млекопитающих. Так же как карбоангидраза, этот фермент содержит один ион Zn2+ на одну белковую макромолекулу, молекулярная масса которой 34300. Биологическая роль карбо-ксипептидазы заключается в катализе процесса гидролиза пептидов - разрыве пептидной связи на карбоксильном конце пептидной цепи:
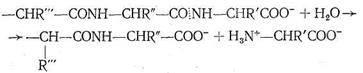
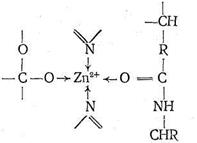
Ион Zn2+ в карбоксипептидазе, так же как в карбоангидразе, находится в полости активного центра фермента, но только два места в его координационной сфере заняты азотом имидазольных остатков гистидина. Третье место занято кислородом другой аминокислоты - глутамина. Четвертое координационное место в координационной сфере Zn2+ принадлежит молекуле воды. Таким образом, и в этом ферменте координационная сфера Zn2+ несимметрична, что, по-видимому, обусловливает напряженность структуры фермента и является обязательным условием каталитической активности комплексных соединений.
|
Предполагают, что одной из важных стадий каталитического процесса, протекающего с участием карбоксипептидазы, является координация ионом Zn2+ атома кислорода карбонила пептидной цепи с вытеснением Н2О, занимающей одно место в координационной сфере. Координация цинком карбонильного кислорода ослабляет связь С-N в пептидной цепи, что облегчает ее разрыв, сопровождающийся гидролизом. Гидролиз разорванной цепи приводит к ослаблению координации карбонильного кислорода пептидной цепи ионом Zn2+. Происходит вытеснение молекулой Н2О, затем вновь следует координация ионом Zn2+ карбонильного кислорода неразорванной цепи и т, д, Ферментативная активность карбоксипептидазы сохраняется, если Zn2+ заменить на ион Со2+, так же как в случае карбоангидразы. По-видимому, это связано с очень близкими величинами констант устойчивости (т. е. сходными термодинамическими, характеристиками) аналогичных комплексов Zn2+ и Со2+, а также одинаковыми размерами этих ионов. Замена Zn2+ в карбоксипептидазе на Gd2+, Hg2+, Cu2+, Pb2+ приводит к исчезновению каталитической активности фермента.
К группе ферментов, катализирующих процессы гидролиза, относятся также киназы, ускоряющие перемещение различных фрагментов биолигандов (например, ионы РО3-)от одного биополимера к другому. Все киназы содержат ионы М2+. Аминопептидазы (ионы Mg2+, Zn2+, Mn2+, Ca2+-кофермент) катализируют гидролиз пептидов, при этом каждый из большого числа ферментов, входящих в эту группу, ускоряет разрыв пептидной связи именно для данного сочетания разноименных аминокислот в пептидной цепи. Фосфатазы катализируют гидролиз сложных эфиров на основе ортофосфорной кислоты.
Ферментативный катализ окислительно-восстановительных реакций
Окислительные реакции, протекающие в живом организме с участием молекулярного кислорода, вносят основной вклад в накопление организмом энергии. Окислительно-восстановительные реакции в отсутствие катализатора всегда протекают медленнее, чем реакции обмена ионов (тоже не катализируемые), например реакции гидролиза. Поэтому роль ферментативного катализа в ускорении окислительно-восстановительных процессов, протекающих в живых организмах, является особенно важной. Окислительно-восстановительные реакции в организме катализируются ферментами, содержащими ионы цинка, железа, меди, молибдена, кобальта. Роль металлсодержащих групп в ферментах, катализирующих окислительно-восстановительные процессы, изучена недостаточно. Однако ясно, что ион металла в ферменте не всегда входит в активный его центр. В ряде случаев ионы металла определяют лишь третичную и четвертичную структуру белка, образующего апофермент, а сам по себе кофермент ионов металла не содержит. Тем не менее роль металла остается крайне важной - замена ионов одного металла на другой меняет структуру фермента и его активность.
Из металлсодержащих ферментов, катализирующих окислительно-восстановительные процессы, наиболее изучены цинксодержащие дегидрогеназы, а также железосодержащие ферменты.
К числу ферментов, катализирующих окислительно-восстановительные реакции, относятся гемсодержащие ферменты - цитохромы. Атомы железа в цитохромах, так же как в гемоглобине и миоглобине, координируют пять атомов азота (порфирина и гистидина), шестое координационное место занимает атом серы аминокислоты - метионина. Известно 50 видов ферментов этого типа, несколько различающихся по составу органической его части. Железо в цитохромах играет роль переносчика электронов - оно принимает электроны от восстановителя и передает их окислителю. Окислителем может быть и кислород, но он не участвует в координации железа, входящего в состав цитохромов, поскольку железо в цитохроме имеет полностью насыщенную координационную сферу.
Среди гемсодержащих ферментов, катализирующих окислительно-восстановительные реакции, также большое значение имеют гем-белки - каталаза и пероксидаза, ускоряющие распад перекиси водорода. Последняя образуется в результате катализируемого ферментами взаимодействия восстановленных форм субстратов с молекулярным кислородом.
Кроме гемсодержащих ферментов известно большое число металлсодержащих ферментов, имеющих «негемовое» строение. К их числу принадлежат ферредоксины, играющие в живых организмах, подобно цитохромам, роль переносчиков электронов. Ферредоксины имеют молекулярную массу от 6000 до 12 000. Атомы железа в них окружены четырьмя атомами серы:
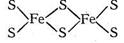
Концевые атомы серы принадлежат цистеиновому фрагменту белка. Считают, что важную роль источника (или «ловушки») электронов играет группировка Fe4S4, содержащаяся в каждом из ферредоксиновых белков.
Окислительно-восстановительные реакции катализируются также металлоферментами, включающими медь и другие металлы. Важное значение имеет оксидаза аскорбиновой кислоты, содержащаяся в растениях и микроорганизмах. Молекулярная масса оксидазы 140 000, на одну такую белковую глобулу приходится восемь атомов меди. Как показывает название, оксидаза катализирует переход аскорбиновой кислоты в дегидроаскорбиновую кислоту. Медь также содержится в белке-гемоцианине, который способен связывать молекулярный кислород (О2 на два атома меди). Используется гемоцианин для транспорта кислорода в организмах низших животных.
К числу ферментов, катализирующих перенос электронов, т. е, окислительно-восстановительные процессы, относится нитрогеназа, содержащаяся в азотистых бактериях. Нитрогеназа катализирует связывание атмосферного азота. С использованием меченого азота (15N2) показано, что бактерии восстанавливают N2 в первую очередь до аммиака. Этот процесс происходит только в присутствии Mo, Fe и Mg. Установлено, что нитрогеназа состоит из двух белков (молекулярная масса 250 000 и 70 000), каждый из которых по отдельности не активен.
Только в совместном присутствии эти белки проявляют каталитический эффект. В одном из белков содержится на одну глобулу один-два атома Мо, 15 атомов Fe, большое количество серы; во втором-молибдена нет. имеются два атома железа и два атома серы (неорганической, не входящей в состав белка).
Хлорофилл в зеленых растениях играет важнейшую роль, связанную с энергетическим обеспечением окислительно-восстановительных процессов при фотосинтезе. Установлено, что хлорофилл, т. е. порфириновый комплекс Mg2+, содержащийся в хлоропласте зеленых частей растения, поглощает световую энергию. Длина волны поглощаемого света (~ 700 нм) определяется системой сопряженных связей в порфириновом комплексе. Вхождение Mg2+ в порфириновый комплекс делает лиганд более жестким, что уменьшает рассеяние энергии в результате колебательных движений.
Световая энергия, запасенная хлорофиллом, расходуется на протекание эндотермической реакции, которая называется реакцией фотосинтеза:
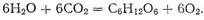
Механизм фотосинтеза в полной мере не изучен. Известно только, что в стадиях фотосинтеза принимают участие комплексы четырех металлов (включая магний хлорофилла). Так, установлено, что в стадии переноса электрона, завершающейся отщеплением молекулярного кислорода, участвует комплекс марганца; на последующих стадиях в реакции вступают комплексы железа (ферредоксин, цитохромы) и меди (пластоцианины).
Таким образом, роль хелатных комплексов металлов в регулировке сложнейших процессов, протекающих в живых и растительных организмах, чрезвычайно важна и многообразна. Отметим еще некоторые из биологически важных процессов, связанных с участием неорганических соединений или ионов.
Другие биологические функции неорганических соединений.
Роль ионов Са2+ в организмах животных и растений сложна. Больше всего кальция содержится в костной ткани. Кристаллы кости имеют приблизительный состав Са10(Р04)6(ОН)2 и, таким образом, относятся к числу гидроксилапатитов. Минеральная часть кости включает, кроме того, карбонаты, фториды, цитраты и гидроокиси металлов, среди которых, кроме Са2+, есть Mg2+, Na+ и К+. Неорганическая часть кости составляет только 0.25 ее массы - остальное органические компоненты.
Среди наиболее важных функций Са2+ - его роль в ферментативных системах, в том числе как регулятора сокращения мышц, передатчика нервного импульса, а также в системе свертывания крови.
В организм кальций вводится в виде среднего фосфата, содержащегося в пище. В пищеварительном тракте под влиянием кислой среды средний фосфат преобразуется в хорошо растворимые кислые фосфаты СаНРО4 и Са(Н2РО4)2. Именно кислые фосфаты всасываются в кишечнике и переходят в плазму крови.
Концентрация ионов Са2+ в крови человека составляет обычно 0,0022-0,0028 моль/л. Примерно половина кальция находится в виде акваионов, способных проходить через мембраны. Другая часть связана с белком (альбумин) и через мембраны не проходит. Интересно, что концентрацию кальция можно определить, используя биологический тест - по частоте сокращения сердца лягушки (или черепахи). Этот способ определения концентрации незакомплексованного Са2+ был использован для определения Куст хелатных комплексов кальция в неорганических исследованиях.
Ионы Са2+ наряду с ионами К+ и Mg2+ влияют не только на частоту сокращения мышц, в том числе сердечной мышцы, но и на действие сердечных гликозидов (типа наперстянки шерстистой - Digitalis). Известно, что при передозировке гликозидов сердце останавливается. Введение при этом ионов К+ и Mg2+ в мышцу сердца ослабляет действие гликозидов, а введение Са2+ - усиливает. Однако ионы Са2+ можно связать в прочный комплекс, например с ЭДТА. Так, если вовремя ввести ЭДТА в мышцу остановившегося сердца, оно вновь начинает биться.
Переизбыток Са2+ оказывает нежелательные воздействия на организм - происходит «образование камней», «отложение солей» и т. д. Так как ионы Са2+ и Mg2+ входят в состав ткани стенок бактериальных клеток, изменение содержания ионов Са2+ в системе может привести к гибели микроорганизма. Такой эффект наблюдается, в частности, если в систему ввести ЭДТА или другой комплексон высокой дентатности. ЭДТА, связывая Mg2+ и Са2+ в прочный комплекс, разрушает стенки бактериальных клеток, что и приводит к гибели микроорганизмов. Вымывание из организма ионов Са2+ и других полезных ионов происходит при использовании комплексонов и хелатообразователей для удаления из организма ионов токсичных металлов, таких, как Hg2+, Pb2+ и др. Чтобы уменьшить вымывание ионов Са2+ при лечении тех или иных заболеваний, в организм вводят ЭДТА в виде кальциевого комплекса СаЭДТА2-.
Ионы щелочных металлов, как уже упоминалось, выполняют в живых и растительных организмах многообразные функции. Это относится не только к ионам Na+ и К+. Установлено, что прием препаратов, содержащих ионы Li+ и Rb+, облегчает состояние больных, у которых обнаружен маниакально-депрессивный синдром.
Передозировка в организме ионов металлов, принадлежащих к числу «металлов жизни», а также случайное введение ионов других металлов (например, Hg2+, Be2+, Cd2+, Pb2+) вызывают тяжелые нарушения жизнедеятельности организма. В эт,их случаях лечение проводят, используя медикаменты (детоксиканты), проявляющие свойства лигандов специфического действия.
Предложено использовать для выведения из организма ионов металлов, проявляющих токсический эффект, следующие лиганды.
Этилендиаминтетраацетат (ЭДТА) - в виде соли кальция - выводит из организма Pb2+, V(IV), V(V). Если ЭДТА применяют дли регулировки свертывания крови, то вводят натриевую соль, чтобы понизить [Са2+].
Ауринтрикарбоксилат или салицилат (натрия) вводят в организм для закомплексовывания бериллия. Комплексы Ве2+ с этими лигандами проходят через биомембраны, и таким образом Ве2+ может быть удален из организма.
D-пеннициламин (HSC(CH3)2-CHNH2-COOH) используют для специфического связывания Сu2+ и выведения меди из организма, в частности, если в результате передозировки или неправильного функционирования организма в печени, мозге и почках накапливается медь (болезнь Вильсона, лейкемия и др.).
Димеркаптал СН2ОН-CHSH-CH2SH закомплексовывает и выводит из организма Hg, As, Те, Tl, Au.
Ферриоксимин-В-полигидроксамовая кислота используется для специфического связывания и выведения из организма избытка железа при заболевании «сидерозис».
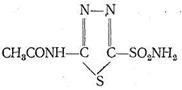
Многие лекарства, применяемые в медицинской практике, представляют собой лиганды, ингибирующие действие активных центров металлов-ферментов путем координационного насыщения ионов металлов. Например, введение в организм диакарба приводит к подавлению ферментативной активности карбоангидразы, катализирующей реакцию связывания воды и СО2в бикарбонат-ион.
|
Ингибирование карбоангидразы диакарбом достигается вследствие координации ионом Zn2+ атома серы диакарба. В результате координация Н2О и СО2 цинком прекращается, вода не связывается в НСО3-я выводится из организма - достигается диуретический эффект. К той же группе лекарств относится дисульфурам который блокирует ионы Сu2+ в ферменте, катализирующем окисление ацетальдегида.
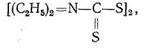
|
Это лекарство используется при лечении алкоголизма, поскольку накопление ацетальдегида в организме вызывает неприятные ощущения и отвращение к алкоголю.
Многие яды действуют по аналогичному механизму. Например, угарный газ и цианиды блокируют ионы железа в гемоглобине и делают невозможным перенос О2 от легких к периферийным тканям. Организм «обескислороживается» и гибнет.
Действие многих лекарств основано на способности комплексных соединений ионов металлов проходить через биомембраны, тогда как акваионы и лиганды, взятые в отдельности, такой способностью обладают в очень малой степени либо вообще не обладают. Примером могут быть антибиотики, активность которых существенно возрастает в присутствии ионов металлов. То же относится к противогрибковым препаратам типа 8-оксихинолината Fe (III). Установлено, что только совместное присутствие акваионов Fe3+*aq и 8-оксихинолина дает антигрибковый эффект. Очевидно, что образующийся комплекс железа (III) проходит через стенки клеток грибков и вызывает их гибель.
Интересные данные получены в последнее время о катионных комплексах типа Co(NH3)63+, Co(NH3)5O22+ и комплексах железа, рутения и других переходных металлов с нейтральными лигандами типа фенантролина и дипиридила. Оказалось, что эти (вероятно, и другие) катионные комплексы переходных металлов сходны по геометрии и плотности положительного заряда с «головкой» ацетилхолина [(CH3)3N+](CH2)2COOCH3. Ацетилхолин является действующим началом яда кураре, вызывающего паралич нервных окончаний. Указанные катионные комплексы вызывают подобное действие, об этом следует помнить, когда в практикумах по неорганической, аналитической я координационной химии проводится синтез такого рода комплексных соединений.
Изучение биологической активности неорганических соединений только начинается. Интерес к испытанию их биологической активности резко возрос после открытия в 1969 г. противораковой активности соли Пейроне - цис-дихлородиамминплатины(Н). Было замечено, что электролиз раствора NH4CI с использованием платиновых электродов приводит к потере способности воспроизводства у кишечной палочки Е. Соli. Это отнесли к положительному действию соли Пейроне, присутствующеи в растворе. Поскольку антираковое и антимикробное действие симбатны, соль Пейроне была испытана на антираковое действие и показала высокую эффективность. Сейчас проводятся поиски столь же эффективных, но менее токсичных медикаментов. Установлено, что антираковое действие зависит от строения комплексов платины (и других металлов). В частности, транс-изомер дихлородиамминплатины(П) не проявляет антиракового действия. Замена атомов С1 на Вr и I делает неактивными и цис-изомер.
Бионеорганическая химия и охрана окружающей среды
Бионеорганические аспекты экологии имеют большое значение. В частности, внедрение в химическую технологию и сельское хозяйство реагентов, способных закомплексовывать ионы металлов, вызывает нежелательные сдвиги природных равновесий. Можно упомянуть, например, действие комплексонов, сбрасываемых в больших количествах в природные водоемы предприятиями, применяющими эти реагенты для «умягчения» воды: фотопромышленностью, красильной (текстильной и полиграфической) промышленностью, энергетикой (введение в воду паровых потоков и отопительной сети комплексонов для предотвращения осадкообразования), нефтяной и цементной промышленностью (предотвращение солеотложений в трубах, замедление схватывания бетона), сельским хозяйством (для борьбы с хлорозом и анемией вводят железо в почву или в пищу животных в виде комплексонатов, не подвергающихся гидролитическому разрушению и легко усвояемому животными, в отличие от акваионов Fe (II), Fe(III) в растворах простых солей). Комплексоны, попадая в природные водоемы, вызывают растворение осадков токсичных металлов, десятилетиями накапливающихся на дне морей и океанов. Переходя в раствор в виде комплексонатов, ионы Hg2+, Zn2+, Cd2+, Pb2+ и других токсичных металлов проникают через биомембраны и отравляют живые организмы. Присутствие комплексонатов в природных водах вызывает гипоксию (недостаток кислорода), а вследствие этого гибель планктона и в конце концов гибель высших животных, стоящих в конце экологической цепи. Кислород тратится на окисление ионов металлов, например на переведение Fe(II) в Ре(III). Комплексоны этому способствуют, так как стабилизируют высшие степени окисления металлов, которым отвечают наиболее, устойчивые комплексы. Поэтому нельзя не отметить важность работ, посвященных синтезу, исследованию и разработке методов использования комплексообразующих агентов, которые легко бы обезвреживались, попадая в условия сброса. Интерес, на наш взгляд, представляют комплексоны типа этилендиаминдиянтарной кислоты. Эти соединения, обладая высокой комплексообразующей способностью, в природных средах быстро дезактивируются и не смещают установившегося равновесия.
В плане охраны окружающей среды могут рассматриваться и работы по моделированию процесса связывания атмосферного азота. Замена энергоемкого производства синтезом азотсодержащих соединений в мягких условиях (подобно условиям связывания N2 азотистыми бактериями) приведет в конечном итоге к сбережению природных ресурсов и охране биосферы.