Раздражимость
Раздражимостью называется способность живых организмов и образующих их систем (органов, тканей, клеток) реагировать на внешнее воздействие изменением своих физико-химических и физиологических свойств.
Раздражимость проявляется в изменениях текущих значений физиологических параметров, величина которых превышает их сдвиги при покое. Раздражимость является универсальным проявлением жизнедеятельности всех без исключения биологических систем.
Возбудимость
Когда изменения внешней среды начинают превышать известный индивидуальный уровень, активное состояние некоторых тканей и клеток может сопровождаться проявлением специфической функции данной живой системы — возбудимостью.
Возбудимостью называется способность организма, органа, ткани или клетки отвечать на раздражение активной специфической реакцией — возбуждением (генерацией нервного импульса, сокращением, секрецией и др.).
Раздражимость и возбудимость характеризуют в сущности одно и то же свойство биологической системы — способность отвечать на внешние воздействия. Однако термин возбудимость используется для определения специфических реакций, имеющих более позднее филогенетическое происхождение. Возбудимость является, следовательно, высшим проявлением более общего свойства раздражимости тканей.
2. Не́рвнаясисте́ма — целостная морфологическая и функциональная совокупность различных взаимосвязанных нервных структур, которая совместно с гуморальной системой обеспечивает взаимосвязанную регуляцию деятельности всех систем организма и реакцию на изменение условий внутренней и внешней среды. Нервная система действует как интегративная система, связывая в одно целое чувствительность, двигательную активность и работу других регуляторных систем (эндокринной и иммунной).
Общая характеристика нервной системы
Все разнообразие значений нервной системы вытекает из ее свойств.
1. Возбудимость, раздражимость и проводимость характеризуются как функции времени, то есть это процесс, возникающий от раздражения до проявления ответной деятельности органа. Согласно электрической теории распространения нервного импульса в нервном волокне он распространяется за счет перехода локальных очагов возбуждения на соседние неактивные области нервного волокна или процесса распространяющейся деполяризации потенциала действия, представляющего подобие электрического тока. В синапсах протекает другой-химический процесс, при котором развитие волны возбуждения-поляризации принадлежит медиатору ацетилхолину, то есть химической реакции.
2. Нервная система обладает свойством трансформации и генерации энергий внешней и внутренней среды и преобразования их в нервный процесс.
3. К особенно важному свойству нервной системы относится свойство мозга хранить информацию в процессе не только онто-, но и филогенеза.
Нейроны
Нервная система состоит из нейронов, или нервных клеток и нейроглии, или нейроглиальных клеток. Нейроны — это основные структурные и функциональные элементы как в центральной, так и периферической нервной системе. Нейроны — это возбудимые клетки, то есть они способны генерировать и передавать электрические импульсы (потенциалы действия). Нейроны имеют различную форму и размеры, формируют отростки двух типов: аксоны и дендриты. У нейрона обычно несколько коротких разветвлённых дендритов, по которым импульсы следуют к телу нейрона, и один длинный аксон, по которому импульсы идут от тела нейрона к другим клеткам (нейронам, мышечным либо железистым клеткам). Передача возбуждения с одного нейрона на другие клетки происходит посредством специализированных контактов — синапсов.
Нейроглия
Глиальные клетки более многочисленны, чем нейроны и составляют по крайней мере половину объёма ЦНС, но в отличие от нейронов они не могут генерировать потенциалов действия. Нейроглиальные клетки различны по строению и происхождению, они выполняют вспомогательные функции в нервной системе, обеспечивая опорную, трофическую, секреторную, разграничительную и защитную функции.
5.
| Нервные волокна - отростки нервных клеток (нейронов), имеющие оболочку и способные проводить нервный импульс. Главной составной частью нервного волокна является отросток нейрона, образующий как бы ось волокна. Большей частью это аксон. Нервный отросток окружен оболочкой сложного строения, вместе с которой он и образует волокно. Толщина нервного волокна в организме человека, как правило, не превышает 30 микрометров. Нервные волокна делятся на мякотные (миелиновые) и безмякотные (безмиелиновые). Первые имеют миелиновую оболочку, покрывающую аксон, вторые лишены миелиновой оболочки. Как в периферической так и в центральной нервной |
| системе преобладают миелиновые волокна. Нервные волокна, лишенные миелина располагаются преимущественно в симпатическом отделе вегетативной нервной системы. В месте отхождения нервного волокна от клетки и в области перехода его в конечные разветвления нервные волокна могут быть лишены всяких оболочек, и тогда они называются голыми осевыми цилиндрами. В зависимости от характера проводимого по ним сигнала, нервные волокна подразделяют на двигательные вегетативные, чувствительные и двигательные соматические. Строение нервных волокон: Миелиновое нервное волокно имеет в своём составе следующие элементы (структуры): 1) осевой цилиндр, располагающийся в самом центре нервного волокна, 2) миелиновую оболочку, покрывающую осевой цилиндр, 3) шванновскую оболочку. Осевой цилиндр состоит из нейрофибрилл. Мякотная оболочка содержит большое количество веществ липоидного характера, известных под названием миелина. Миелин обеспечивает быстроту проведение нервных импульсов. Миелиновая оболочка покрывает осевой цилиндр не на всём промежутке, образуя промежутки, получившие название перехваты Ранвье. В области перехватов Ранвье осевой цилиндр нервного волокна примыкает к верхней - шванновской оболочке. Промежуток волокна, расположенный между двумя перехватами Ранвье, называют сегментом волокна. В каждом таком сегменте на окрашенных препаратах можно видеть ядро шванновской оболочки. Оно лежит приблизительно посредине сегмента и окружено протоплазмой шванновской клетки, в петлях которой и содержится миелин. Между перехватами Ранвье миелиновая оболочка также не является сплошной. В толще ее обнаруживаются так называемые насечки Шмидт-Лантермана, идущие в косом направлении. Клетки шванновской оболочки, так же как и нейроны с отростками, развиваются из эктодермы. Они покрывают осевой цилиндр нервного волокна периферической нервной системы аналогично тому, как клетки глии покрывают нервное волокно в центральной нервной системе. В результате этого они могут называться периферическими глиальными клетками. В центральной нервной системе нервные волокна не имеют шванновских оболочек. Роль шванновских клеток здесь выполняют элементы олигодендроглии. Безмиелиновое (безмякотное) нервное волокно лишено миелиновой обкладки и состоит только из осевого цилиндра и шванновской оболочки. Функция нервных волокон. Главная функция нервных волокон – передача нервного импульса. В настоящее время изучено два типа нервной передачи: импульсная и безимпульсная. Импульсная передача обеспечивается электролитными и нейротрансмиттерными механизмами. Скорость передачи нервного импульса в миелиновых волокнах значительно выше, чем в безмякотных. В её осуществлении важнейшая роль принадлежит миелину. Данное вещество способно изолировать нервный импульс, в результате чего передача сигнала по нервному волокну происходит скачкообразно, от одного перехвата Ранвье к другому. Безимпульсная передача осуществляется током аксоплазмы по специальным микротрубочкам аксона, содержащим трофогены – вещества, оказывающие на иннервируемый орган трофическое влияние. |
6. Моторная бляшка. Электронная микрофотограмма. 33 000.
1 - концевые веточки нервного волокна; 2 - митохондрии в аксоплазме; 3 - синаптические пузырьки в аксоплазме; 4 - аксолемма, образующая в этом месте пресинаптическую мембрану; 5 - сарколемма, образующая в этом месте постсинаптическую мембрану; 6 - складки постсинаптической мембраны; 7 - синаптическая щель; 8 - леммоцит (шванновская клетка); 9 - саркоплазма и 10 - ядро мышечного волокна (по Г. Еляковой).
КОММЕНТАРИИ К ЭЛЕКТРОНОГРАММЕ
Моторная бляшка (нейромышечный синапс) - эффекторное (двигательное) нервное окончание, кото-рое встречается только в скелетной мышечной ткани. Нервное волокно (аксон + леммоцит) контактирует с миосимпластом.
Строение моторной бляшки похоже на строение классического синапса в нервной ткани. Поэтому видны следующие структуры:
1. Леммоцит (шванновская клетка) (цифра 8) - "прикрывает контакт сверху, изолируя и защищая его. В его цитоплазме видны митохондрии и цистерны гранулярной ЭПС.
2. Аксон двигательного нейрона (из передних рогов спинного мозга) (цифра 1), возле моторной бляшки уже не имеет миелиновой оболочки. Его аксолемма (цитолемма) (цифра 4) выполняет рольпресинаптической части синапса, поэтому в его аксоплазме много синаптических пузырьков (цифра 3), содержащих ацетилхолин (является медиатором в моторной бляшке). Кроме того имеются митохондрии, обеспечивающие энергию для транспорта медиатора из тела нейрона и его обратного захвата из синаптической щели.
3. Миосимпласт (мышечное волокна) в области моторной бляшки теряет поперечную исчерченность. В данном случае видны одно из его многочисленных ядер (цифра 10) и саркоплазма (цифра 9) - его сарколемма (цифра 5) выполняет роль постсинаптической мембраны и образует многочисленные складки в области синапса, для увеличения площади контакта с медиатором (цифра 6).
4. Синаптическая щель (цифра 7).
КРАТКАЯ ИНФОРМАЦИЯ
Аксон мотонейрона выделяет ацетилхолин в синаптическую щель ==> рецепторы на постсинаптической мембране (в данном случае - сарколемме) возбуждаются ==> происходит деполяризация сарколеммы ==> волна деполяризации заходит в Т-трубочки (поперечные впячивания сарколеммы). Т-трубочки объединены в триады с концевыми цистернами (последние - это части саркоплазматического ретикулума, где депонируется Са++), следовательно, ==> возбуждение передается и на ретикулум, кальций выходит в саркоплазму ==> связывается с тонкими нитями, освобождая их активные центра для контакта с актином. Начинается сокращение миосимпласта.
Когда импульс перестает "приходить" по аксону, оставшийся в синаптической щели ацетилхолин разрушается ферментом ацетилхолинэстеразой или захватывается аксоном.
В скелетной мышечной ткани каждый симпласт имеет нейромышечный синапс, в отличие от гладкой и сердечной мышечных тканей, где один аксон просто ветвится среди 8-10 миоцитов. Нервное окончание в этом случае простое, свободное. Возбуждение между миоцитами передается по нексусам.
1 миосимпласт + 1 нервное волокно, образующее вним синапс + капилляры вокруг симпласта (в составе эндонев-рия) = мион.
7. Рецепторные (чувствительные) нервные окончания
Эти нервые окончания - рецепторы - рассеяны по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов. Соответственно выделяют две большие группы рецепторов:
· Экстерорецепторы - стимулируемые окружающей средой
o контактцепторы, воспринимающие раздражения, наносимые извне, и падающие непосредственно на ткани организма (болевые, температурные, тактильные и др.)
o дистантцепторы, воспринимающие раздражения от источников, которые находятся на расстоянии (свет, звук)
· Интерорецепторы
o проприоцепторы, воспринимающие раздражения, возникающие внутри организма, в его глубоких тканях, связанных с функцией сохранения положения тела при движениях. Данный вид рецепторов представлен в мышцах, сухожилиях, связках, суставах, надкостнице, импульсы возникают в связи с изменением степени натяжения сухожилий, напряжения мышц и ориентируют в отношении положения тела и частей его в пространстве: отсюда еще наименование - "суставно-мышечное чувство", или "чувство положения и движения (кинестетическое чувство)".
o висцерорецепторы воспринимающие раздражения от внутренних органов. Обычно от данных рецепторов информация очень редко доходит до сознания, например информация от барорецепторов, расположенных в каротидном синусе, которые непрерывно контролируют артериальное давление
В клинике приобрела довольно широкое распространение другая классификация, основанная на биологических данных [показать].
С точки зрения биологических данных чувствительные нервные окончания рассматривается как соотношение и взаимодействие двух систем.
Одна, более древняя, свойственная более примитивной нервной системе, служит для проведения и восприятия сильных, резких, угрожающих целости организма раздражений; сюда относятся грубые болевые и температурные раздражения, связанные с древним "чувствующим" органом - зрительным бугром. Данная система чувствительности носит название протопатической, витальной, ноцицептивноп, таламической.
Другая система связана всецело с корой головного мозга. Являясь более новой и совершенной, она служит для тонкого распознавания качества, характера, степени и локализации раздражения. Сюда относятся такие виды чувствительности, как осязание, определение положения и движения, формы, места нанесения раздражения, различение тонких температурных колебаний, качества боли и т.д. Наименование этой системы чувствительности - эпикритическая, гностическая, корковая.
Эпикритическая чувствительность как система более новая, корковая, якобы оказывает тормозящее влияние на старинную протопатическую подкорковую чувствительность. Предполагалось, что в норме чувствительная функция человека определяется сосуществованием обеих систем в их определенной взаимосвязи; при этом эпикритическая чувствительность вносит элементы точного различения и анализа.
Такое подразделение чувствительности на два отдельных вида вызывает ряд серьезных возражений. Малодоказательным является представление об их соотношении как низшей и высшей систем, о тормозной функции эпикритической по отношению к протопатической; трудно представить себе роль зрительного бугра как органа, "воспринимающего" отдельные виды чувствительности.
В целостном организме любой вид чувствительности связан с работой коры больших полушарий, ибо всякое ощущение как акт сознания немыслимо без участия высших отделов головного мозга. Вместе с тем не подлежит сомнению, что в сложной чувствительности человека, достигшей высокого совершенства в процессе развития, существует представительство и древних примитивных систем, связанных с действием подкорковых, стволовых, сегментарных аппаратов. При поражении, выключении одного из звеньев высокодифференцированной чувствительной системы, значение в которой зрительного бугра все же остается несомненным, мы получаем качественно совершенно иную функциональную систему со своеобразным извращением ощущений и восприятий.
Более употребительной в клинической практике является описательная классификация, основанная на различении вида раздражения и возникающего в связи с ним ощушения.
В зависимости от специфичности раздражения, воспринимаемого данным видом рецептора, все чувствительные окончания разделяют на

· механорецепторы (рис.2)
o медленно адаптирующиеся - например, давление веса тела на подошву. К ним относятся диск Меркеля - реагируют на деформацию перпендикулярно поверхности кожи, окончания Руффини (в безволосой коже) - реагируют на растяжение. В коже, покрытой волосами, диски Меркеля группируются под кожными возвышениями - тельцами Пинкуса-Игго.
o быстро адаптирующиеся - реагирует только на механические стимулы, которые изменяются во времени. К ним относится тельце Мейснера (в безволосой коже), рецептор волосяного фолликула (в оволосненной коже)
o очень быстро адаптирующиеся - реагируют на изменение скорости механической стимуляции. К ним относятся тельца Пачини. Тельца Пачини можно также назвать рецептором вибрации.
· терморецепторы - температурная чувствительность (чувство холода и чувство тепла) и бессознательная регуляция температуры тела
· барорецепторы - чувствительность к изменению артериального давления
· хеморецепторы - чувствительность к понижению парциального давления кислорода и повышению углекислого газа, регулируют дыхание
· ноцицепторы - чувство боли (поверхностной, глубокой, от внутренних органов)
· рецепторы чувства позы, движения, мышечного усилия и др.
По особенностям строения чувствительные окончания подразделяются на
· свободные нервные окончания, т. е. состоящие только из конечных ветвлений осевого цилиндра. Характерны для эпителия (холодовые рецепторы).
В этом случае миелиновые нервные волокна подходят к эпителиальному пласту, теряют миелин, а осевые цилиндры проникают в эпителий и распадаются там между клетками на тонкие терминальные ветви. В многослойном эпителии есть окончания, в состав которых входят, кроме терминалей отростков нервных клеток, специфически измененные эпителиальные клетки - осязательные эпителиоциты. От других клеток эпителия они отличаются светлой цитоплазмой, наличием осмиофильных гранул диаметром 65-180 нм и уплощенным темным ядром. Концевые нервные веточки подходят к таким клеткам и расширяются, образуя дисковидные концевые структуры, связанные с основаниями осязательныхэпителиоцитов.
· несвободные нервные окончания - содержащие в своем составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и клетки глии
o неинкапсулированныe - не имеющие соединительнотканной капсулы
o инкапсулированныe - покрыты соединительнотканной капсулой. К ним относят пластинчатые тельца - рецепторы соединительной ткани, воспринимающие давление; осязательные тельца, находящиеся в составе сосочков кожи (тепловые рецепторы); мышечные веретена - рецепторы скелетных мышц и сухожилий, регистрирующие изменения длины мышечных волокон и скорость этих изменений; нервно-сухожильные веретена - так же рецепторы скелетных мышц и сухожилий, реагирующие на напряжение, прилагаемое к сухожилию при сокращении мышц.
Веретено состоит из нескольких (до 10-12) тонких и коротких поперечнополосатых мышечных волокон, заключенных в растяжимую соединительнотканную капсулу, - интрафузальных волокон.
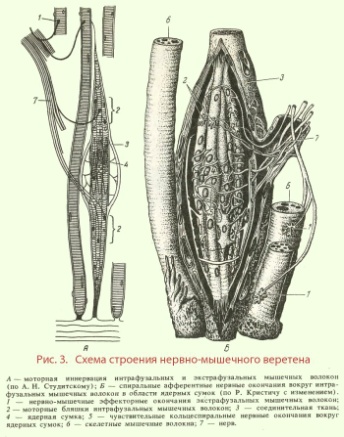
Остальные волокна мышцы лежат за пределами капсулы и называются экстрафузальными (рис. 3, А, Б). Интрафузальные волокна имеют актиновые и миозиновыемиофиламенты только на концах, которые и сокращаются. Рецепторной частью интрафузального мышечного волокна является центральная, несокращающаяся часть.
Различают интрафузальные волокна двух типов: волокна с ядерной сумкой и волокна с ядерной цепочкой.
Волокон с ядерной сумкой в веретене 1-3. В центральной расширенной части они содержат много ядер.
Волокон с ядерной цепочкой в веретене может насчитываться 3-7. Они вдвое тоньше и вдвое короче, чем волокна с ядерной сумкой, и ядра в них расположены цепочкой по всей рецепторной области. К интрафузальным мышечным волокнам подходят афферентные волокна двух типов.
Первичные волокна диаметром 17 мкм образуют окончания в виде спирали - кольцеспиральные окончания как на волокнах с ядерной сумкой, так и на волокнах с ядерной цепочкой.
Вторичные волокна диаметром 8 мкм иннервируют волокна с ядерной цепочкой. По обеим сторонам от кольцеспирального окончания они образуют гроздъевидные окончания.
При расслаблении (или растяжении) мышцы увеличивается и длина интрафузальных волокон, что регистрируется рецепторами. Кольцеспиральные окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, гроздьевидные окончания - только на изменение длины. При внезапном растяжении из кольцеспиральных окончаний в спинной мозг поступает сильный сигнал, который вызывает резкое сокращение мышцы, с которой поступил сигнал - динамический рефлекс на растяжение. При медленном, длительном растяжении волокна возникает статический сигнал на растяжение, передаваемый как от кольцеспиральных, так и от гроздьевидных рецепторов. Этот сигнал может поддерживать мышцу в состоянии сокращения в течение нескольких часов.
Интрафузальные волокна имеют также эфферентную иннервацию. К ним подходят тонкие моторные волокна, оканчивающиеся аксомышечным синапсом на концах мышечного волокна. Вызывая сокращение концевых участков интрафузального волокна, они усиливают растяжение центральной рецепторной его части, повышая реакцию рецептора.
Нервно-сухожильные веретена обычно располагаются в месте соединения мышцы с сухожилием. Коллагеновые пучки сухожилия, связанные с 10-15 мышечными волокнами, окружены соединительнотканной капсулой. К нервно-сухожильному веретену подходит толстое (диаметром около 16 мкм) миелиновое волокно, которое теряет миелин и образует терминали, ветвящиеся между пучками коллагеновых волокон сухожилия. Сигнал с нервно-сухожильных веретен, вызванный напряжением мышцы, возбуждает тормозные нейроны спинного мозга. Последние тормозят соответствующие двигательные нейроны, предотвращая перерастяжение мышцы.
8 билет нейросекреторные клетки
Нейросекреторные клетки являют из себя многоцелевую для дерганых систем многоклеточных животных тип клеточных деталей тканей психики. Отличительной сложности системы этих клеток достигают из числа позвоночных у млекопитающих и у высочайших адептов первичноротых животных. Широко представлены они и у адептов основной массы классов иных типов многоклеточных животных. Случаем стереотипной нейросекреторной клетки имеют все шансы работать клетки гипофизарно-гипоталамической области у млекопитающих и человека (рис. 101). По экстерьеру Нейросекреторные клетки сходны с мультиполярным нейроном. Они имеют некоторое количество коротких дендритов с множественными химическими синаптическими завершениями аксонов дерганых клеток и 1 длинный аксон, заканчивающийся булавовидным расширением, или на сосудах задней доли гипофиза, или в железистой ткани аденогипофиза (рис. 99). Аксоны каких-либо нейросекреторных клеток выделяют коллатерали, по коим нейросекрет имеет возможность транспортироваться к нейронам, иннервирующим нейросекреторные клетки. Данные коллатерали образуют на телах и дендритах нейронов аксосоматические и аксодендрические синапсы, в коих нейрогормоны функционируют как стереотипные медиаторы химических синапсов.
Цитологической спецификой нейросекреторных клеток гипоталамуса считается еще наиболее мощное, нежели в нейроне, становление белок-синтезирующего агрегата (особо у нейросекреторных клеток, продуцирующих пептидныенейрогормоны) и текстур, сознательных за автотранспорт гранул нейросекрета. В химическом отношении Нейросекреторные клетки гипотала-мической области разделяются на 2 немалые категории - пептидэрические и монаминэргические. Первые продуцируют пептидные гормоны, вторые - монаминовые (дофамин, норадреналин, серотонин).
Из числа пептидэргических нейросекреторных клеток гипота-ламической области в биологическом проекте возможно выделить категорию висцеротропных, клеток, гормоны коих работают на висцеральные органы. Они выделяют вазопрессин (антидиуретический гормон), окситоцин и гомологи данных пептидов. Вторая подгруппа нейросекреторных клеток выделяет аденогипо-физтропные гормоны, т. е. гормоны, регулирующие работа железистых клеток аденогипофиза. Из числа выделяемых ими пептидных гормонов наличествуют и гормоны, подстегивающие железистую функцию клеток аденогипофиза - либерины, либо, напротив, угнетающие гормоны - статины. Монаминэргические Нейросекреторные клетки выделяют нейрогормоны как правило в портальную сосудистую систему задней доли гипофиза,
НЕЙРОГОРМОНЫ
(от нейро... и гормоны), биологически активные вещества, вырабатываемые нейросекреторными клетками нервной ткани. Поступают в кровь, гемолимфу, в тканевую или спинномозговую жидкость, оказывают дистантное длительное регулирующее действие. Участвуют в поддержании гомеостаза (в т. ч. водно-солевого), регуляции тонуса гладкой мускулатуры, разл. сторон метаболизма, функций клеток эндокринных желёз и, в целом, в осуществлении защитно-приспособит. реакций организма. Образуются как у беспозвоночных, так и у позвоночных животных. У последних Н. синтезируются нейросекреторными клетками гипоталамуса (окситоцин, вазопрессин и их аналоги, рилизинг-гормоны, дофамин, норадреналин, серотонин), спинного мозга (у рыб — полипептидные Н., уротензины), а также пинеалоцитами эпифиза (серотонин и мелатонин), клетками хромаффинной ткани мозгового вещества надпочечников, параганглиев, ганглиев и нервных стволов периферич. вегетативной нервной системы (норадреналин и адреналин). По химич. природе Н.— полипептиды, катехоламины, 5-ок-ситриптамин (серотонин) и его производное мелатонин. В секреторных гранулах Н., как правило, связаны с белками-носителями (напр., вазопрессин и окситоцин — с нейрофизинами, а катехоламины — с хромогранином)
Нейромодуляторы - химические вещества, которые действуют как нейромедиаторы, но не ограничиваются синаптической щелью, а рассредотачиваются повсюду, модулируя действие многих нейронов в определенной области.
Гистогенез нервной ткани
Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма по средней линии спины дифференцируется и утолщается, формируя нервную пластинку, края которой утолщаются и приподнимаются, образуя нервные валики. Между ними формируется нервный желобок. Нервные валики сближаются и замыкаются в нервную трубку и обособляются от эктодермы. Производными нервной трубки являются нейроны и макроглия центральной нервной системы (ЦНС).
При смыкании нервной трубки в области нервных валиков между ней и эпидермальной эктодермой с обеих сторон выделяются скопления клеток, образующие нервный гребень, называемый также ганглиозной пластинкой. Производными нервного гребня являются: нейроны чувствительных (сенсорных) и автономных ганглиев, клетки мягкой мозговой и паутинной оболочек мозга и некоторые виды глии: нейролеммоциты, клетки-сателлиты ганглиев, клетки мозгового вещества надпочечников, меланоциты кожи, часть клеток диффузной эндокринной системы.
В формировании ганглиев V, VII, IX и Х пар черепных нервов принимают участие нейрональныеплакоды – утолщенные участки эктодермы по бокам от нервной трубки в краниальной части зародыша. Производными плакод также являются некоторые клетки органов чувств – слуха, равновесия, вкуса (рецепторные, поддерживающие и выстилающие канальцы) и зрения (эпителий хрусталика).
Стенка нервной трубки на ранних стадиях развития состоит из одного слоя клеток призматической формы, которые интенсивно делятся и мигрируют от её просвета, в результате на 3-4 неделе в ней можно выделить 4 концентрических зоны (изнутри наружу):
1.
вентрикулярная (матричная, эпиндимная) содержит камбиальные элементы и митотически делящиеся клетки. Матричные клетки являются предшественниками нейронов и клеток макроглии;
2.
субвентрикулярная зона состоит из клеток, утративших способность к перемещению ядер, но сохраняющих высокую пролиферативную активность;
3.
промежуточная (плащевая, мантийная) зона состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон – нейробластов и глиобластов;
4.
маргинальная зона (краевая вуаль) формируется из врастающих в неё аксонов нейробластов и макроглии и дает начало белому веществу.
По мере дифференцировки нейробласта изменяется субмикроскопическое строение его ядра и цитоплазмы. В ядре возникают участки различной электронной плотности в виде мелких зерен и нитей. В цитоплазме происходит накопление цистерн ГрЭС, увеличение объема комплекса Гольджи, уменьшается количество свободных рибосом и полисом. Специфическим признаком начавшейся специализации нервных клеток следует считать появление в их цитоплазме пучков нейрофиламентов и микротрубочек. Тело нейробласта постепенно приобретает грушевидную форму, от заостренного конца начинает развиваться отросток-аксон (нейрит), а позднее – другие отростки (дендриты). Нейробласты превращаются в зрелые нервные клетки и между ними устанавливаются контакты (синапсы).
В процессе дифференцировки нейронов из нейробластов различают домедиаторный и медиаторный периоды.
Значительная часть нейроцитов в ходе гистогенеза нервной ткани погибает (от 20 до 80%) путем апоптоза – запрограммированной физиологической гибели клеток.
При дифференцировке клеток глиогенной линии вначале образуются глиобласты. Изних образуется астроцитарнаяглия и олигодендроглия.
10. У кишечнополостных существуют и типичные синапсы, передача нервных импульсов в которых осуществляется посредством химических медиаторов. Такого рода синапсы могут обладать различной по сложности структурой, а также различным химизмом. Даже у наиболее примитивного кишечнополостного – гидры (Hydravulgaris) обнаружены поляризованные синапсы, содержащие везикулы с электронноплотным содержимым. Однако нервные сети многих кишечнополостных могут достаточно эффективно координировать и общую двигательную, активность этих животных. Возможность такой координации связана с наличием определенной дифференцировки нервной сети, с усложнением взаимодействия различных нервных сетей животного между собой (рис. 5).
В нервных сетях кишечнополостных наряду с неполяризованными, имеются также и поляризованные синапсы. Полярность особого рода создается неодинаковой чувствительностью различных участков тела животного к внешним воздействиям. Исключительно важную роль в механизмах координации играют имеющиеся в нервных сетях кишечнополостных специализированные пути более быстрой передачи нервных импульсов
11У хордовых вся центральная нервная система представлена нервной трубкой, лежащей со спинной стороны животного. Передний конец трубки обыкновенно расширен и образует головной мозг, между тем как задняя цилиндрическая часть трубки получает название спинного мозга. У зародыша центральная нервная система закладывается в виде продольной полосы утолщенной эктодермы, нервной пластинки, которая углубляется, образуя жолоб, замыкающийся затем в трубку. Передний конец трубки остается довольно долго открытым и это отверстие носит название невропора. Задний конец трубки у зародыша сообщается с полостью кишки (нервно-кишечный канал).Закладка центральной нервной системы в виде полосы утолщенной вктодермы наблюдается нередко и у беспозвоночных, но у них она не замыкается в трубку. Расположение нервных элементов у хордовых также иное: нервные клетки помещаются в центральной части органа, а волокна — в периферической.1 Имеются и еще более непосредственные указания иа первоначальную чувствующую функцию нервной трубки: в нижней стенке промежуточного мозга, именно в сосудистом мешке рыб, а также в эпендиме вокруг центрального канала спинного моэга позвоночных также найдены чувствующие клетки.
12У спинальных рыб отмечается рефлекторная активность всего тела, которая прекращается после стимуляции. Для медулярных рыб характерны ритмические движения или всего туловища (вьюны,золотые рыбки), или только плавников (Sargus, Labrus, Serranus). Движения мезэнцефальных рыб еще сложнее. Причина различий заключается в наличии ганглиозных структур в продолговатом и среднем мозге, оказывающих нисходящее активизирующее влияние на спинной мозг. Клетки,лежащие в основании среднего мозга, выполняют функцию, аналогичную функциям красного ядра среднего мозга наземных позвоночных. Эти клетки (nucleusrelicularismesencephali) играют большую роль в регуляции координации движений рыб через распределение тонуса антагонистической мускулатуры. Данное явление характерно только для костистых рыб. У более примитивных рыб, например акул, спинной мозг имеет большую функциональную самостоятельность.
Расчленение головного и спинного мозга у хрящевых рыб не нарушает ритмическую активность спинного мозга при условии, что рыба находится во взвешенном положении. Если хрящевую спинальную рыбу положить на грунт, то она утрачивает ритмическую активность. Другими словами, даже у хрящевых рыб активность спинного мозга находится под влиянием головного мозга(по крайней мере, вестибулярного аппарата). Возникновение сложного сенсорного аппарата и морфофункциональное усложнение опорно-двигательного аппарата предопределили дальнейшуюцефализацию нервной системы костистых рыб. Несмотря на разницу в дифференциации головного мозга в группе костистых рыб, для них характерны общие филогенетические приобретения:расширение связей между головным и туловищным отделом центральной нервной системы. Прежде всего это нисходящие пути, связывающие аппараты центральной регуляции движений со спинным мозгом:вестибулоспинальный тракт, церебеллоспинальный тракт и бульбоспинальный тракт.
13 билет
Хрящевые рыбы
Нервная система современных хрящевых рыб, как хищников, имеет усложненную строение по сравнению с древними костными рыбами. В туловищной области спинномозговые нервы, образованные слиянием в общие сег-ментальные нервы брюшных и спинных корешков, образуют сложные сплетения, иннервирующие парные плавники. Головной мозг имеет прогрессивно организован передний мозг с сводом с нервной вещества. Промежуточный мозг четко расчленен на отделы и мозговую воронку.Средний мозг становится главным центром зрения. Продолговатый мозг служит центром стато-акустики и органов боковой линии, а также всего висцерального аппарата (челюстного и жаберного). Мозжечок, ведающий моторикой всего тела, прогрессивно развит. Главные нервы в количестве десяти пар имеют затруднено разветвления с включением в черепные нервы 12-й пары (подъязычного нерва).
костные рыбы
Центральная нервная система имеет вид трубки. Передняя часть ее видоизменена в головной мозг, защищенный костями черепной коробки. В головном мозге позвоночных животных различают передний мозг, промежуточный мозг, средний мозг, мозжечок, продолговатый мозг. У рыб мозг невелик. Наиболее развиты средний мозг и мозжечок, управляющие равновесием рыбы и координацией ее движения.
Ориентацию рыб в воде обеспечивают органы чувств: зрение, слух, обоняние, осязание, вкус, а также орган особого чувства — боковая линия. Это погруженный в кожу канал, в котором располагаются нервные окончания, воспринимающие изменения давления и направления тока воды. С внешней средой канал сообщается через отверстия в чешуях, покрывающих канал сверху.
Различия
Их отличает строение скелета: у хрящевых рыб он состоит из хряща, у костных - из костной ткани. Видимо, почти все значительные различия между строением хрящеых и костных рыб обусловлены тем, что первые сформировались в открытом океане, а вторые - в прибрежной зоне.
Особености
Нервная система. У рыб она представлена центральной нервной системой и связанной с ней периферической и вегетативной (симпатической) нервной системой.Центральная нервная система состоит из головного и спинного мозга. К периферической нервной системе относятся нервы, отходящие от головного и спинного мозга к органам. Вегетативная нервная сис