В завис-ти от изолированности, регулярности и постоянства связей между особями, выделяют 3 группы поп-ций: 1)Элементарная (локальная) поп-ция – совок-ть особей вида, населяющие небольшой участок однородной площади. Кол-во таких поп-ций пропорциональна разнообразию условий биогеоценоза. Совокупность элементарных поп-ций формируетболее крупную экологическую популяцию. 2. Экологическая поп-ция – совок-ть элементарных поп-ций, т.е то видовые группировки, приуроченные к конкретным биогеоценозов. Экологич. поп-ции практически ничем не отграничены от окружающих территорий и потому наименее устойчивы по составу. Составляющие ее элементарные поп-ции весьма слабо изолированы др. от др., м/ду ними может происходить довольно часто обмен генетич. инф-цией, но существенно реже, нежели между элементарными поп-циями.
Экологич. поп-ции, слагаясь, образуют географические поп-ции. 3. Географическая поп-ция – совок-ть групп особей предыдущего ранга, кот-е заселяют значит.территорию с географич. однородными условиями среды. В рамках последнего наблюдается единый ритм жизненных явлений и др. функциональные особенности, кот-е создают морфо-физиологический тип, кот-й отличает данную поп-цию от соседних, находящихся в др. географич. Условиях.
В зависимости от способов размножения выд. След. Поп-ции:
1)Поп-ция апомиктов - популяция особей, размножающихся путём партеногенеза. Апомиксис – размнож-е орг-мов, не сопровождающееся половым процессом. В более узком понимании апомиксис — вторично бесполое размножение, при котором зародыш развивается без оплодотворения вследствие нарушения предшествующих этапов полового размножения. В зависимости от того, даёт ли начало новому организму половая (лицевая) или вегетативная клетка, различают две основные формы апомиксиса — партеногенез и апогамию. 2)популяция самоопылителей - популяция растений, продуцирующие цветки, способные опыляться оплодотворяться собственной пыльцой, давая при этом вполне жизнеспособное потомство. 3)Популяция перекрёстников- растения, у которых, как правило, опыление рыльца цветка происходит пыльцой с другого цветка того же растения (гейтеногамия) или с другого растения данного вида (ксеногамия).Опыление происходит при помощи ветра (анемофилия), насекомых (энтомофилия), птиц (орнитофилия), воды (гидрофилия).Перекрестники имеют значительное преимущество перед самоопылителями, давая потомство более жизнеспособное, с более широкими границами приспособляемости к условиям существования. 4)Популяции, которые размножаются бесполым путем.
2.Подходы к определению понятия популяция. Чем шире значение к.-либо понятия, тем с большими трудностями встречается его точное определ-е. В то же время по мере развития любой науки ее осн. понятия неизбежно требуют более строгого определ-я. До 40—50-х годов слову «популяция», бытовавшему в науке, начиная с конца XVIII давалось строгого терминологического знач-я, и оно использ-лось в значении «любая группа особей». С развитием популяц-х исследований такое общее обознач-е оказалось недостаточ-м, и с экологич. точки зрения под поп-цией имели в виду, как правило, естественную, пространственно единую группу особей одного вида, как целое реагир-щую на действие различных внеш. факторов форму существов-я вида конкрет. условиях. Для различий между очень крупными и мелкими частями видового насел-я(в равной степени являющим популяциями при таком широком подходе) было предложено неск. десятков прилагательных: «экологическая», «ландшафтная», «графическая», «биотопическая», «псевдопопуляция» и т.д.
2-м направлением развития представлений о поп-ции было генетическое. Популяцион. генетика была первой из оформившихся популяц-ных дисциплин, и многие её формулировки до сих пор широко использ-тся. В генетике под попу-цией понимается опред. Генетич. система, кот-ю в самой общей форме можно определить как находящуюся в динамич. равновесии систему генотипов, т. е. как группу особей, не просто связ-хпосредством скрещив-я, а связанных особым образом так, что они составляют части единого целого.
Для генетич. расчетов, а затем и для математич. моделирования необходима была формализация определ-я поп-ции как генетич. системы. Под менделевской, или идеальной, популяцией стали иметь в виду неограниченно большую группу особей, все члены кот-й имеют равную возможность скрещивания с особями противополож. пола и внутри кот-й не действуют к.-либо эволюц-ные силы, сдвигающие соотнош-е частот генотипов. Таких групп особей в природе не существуют (всегда есть отклон-я от панмиксии и всегда действуют те или иные эволюц-ные факторы).
Для определ-я понятия поп-ция, отраж-го реальн. положение в природе, понятию мендлевской поп-ции было придано знач-е пространственно-временной группы особей, внутри кот-й реально осуществляется скрещ-е, т е. любой группы размнож-хся особей, имеющих общий генофонд.Самой большой из менделевских поп-ций оказывается вид, самой мелкой — группа, состоящая из самца и самки. Но при таком широком определении термин «поп-ция» оказ-тся настолько неопределенным, что его употребл-е явно теряет терминологическое знач-е. Не спасает положения и использ-е понятий —дем и соседство в смысле природной группировки, в пределах кот-й практически осущ. панмиксия для подавляющей части особей. В таком идеализированном виде и эти понятия оказались далекими от реал-х природн. ситуаций.
3-е, ближайшее к нам по времени направл-е развития представлений о поп-ции можно считать синтетическим: оно как генетич-е, так и экологич-е аспекты. При этом основой такого синтеза явл. учение о микроэволюции. Именно в учении о микроэволюциистала особенно ясна роль поп-ции как элемент-ной эволюциной единицы. В наст.время возникла возможность предложить новый вариант определ-я этого понятия.
Поп-ция — это мин. Самовоспроизв-щаяся группа особей одного вида, на протяжении эволюц-но длит-го времени населяющая опред. пространство, образ-щая самостоят. Генетич. систему и формирующая собственное экологич. гиперпространство. Из этого определ-я вытекает, что поп-ция — это всегда достаточно многочис-ная группа особей, на протяжении большого числа поколений в высокой степени изолир-ная от др. аналогич. групп особей.
4.Подходы к изучению популяции. 1)Генетический подход. Заним. особое— основополагающее место в исследов-ях природ-х поп-ций. Это централ-е полож-е опред-тся тем, что устойчивые, эволюционно значимые измен-я любых — экологич-х, морфологич-х, физиологич-х и др. — св-в поп-ции обязательно должны быть связаны с изменением генетич. структуры поп-ции. Главн. задача популяционной генетики — выяснение закономерностей генетич. струк-ры (состава) поп-ций. От др. направлений популяц-й биологии она отлич. методологически: ее интересует не столько признак, сколько его наследственная обусловл-ть и вероятность появл-я (или исчезнов-я) в чреде поколений. 2)Экологический подход. Экология изучает закономер-ти взаимоотнош-я орг-ма и окруж. среды. В популяц-й биологии экологич-й подход означает выяснениесвязей поп-ции как целостной системы (своеобразного организма) с внешними (по отнош-ю к поп-ции) факторами- как биотическими, так и абиотическими. Это означает исслед-е вещественно-энергетич-х и информац-х параметров поп-ции, определ-е параметров экологич-го гиперпространства, формируемого поп-цией. Все это означает выясн-е места поп-ции как консумента, продуцента или редуцента, места разных возрастно-половых групп в трофической паутине конкретных биогеоценозов, а также иных (кроме трофических) связей поп-ции. 3)Морфологический подход. Осн.задача популяц-й морфологии сост. в изучении стр-ры, состава динамики поп-ций посредством выясн-я морфологич-х признаков, т. е. любых поддающихся сравнению структур-х особенностей особей. Несомненно, что морфологич-й подход в изучении поп-ции частично перекрывается с генетич-м, экологич-м, фенетич-м и онтогенетич-м — во всех этих случаях часто исследуют именно морфологич-епризнак. Так любое изучение меланизма в природ-х поп-циях бабочки или хомячка явл. популяционно-морфологическим исследованием.Морфологич-м оказ-тся и изуч-е хромосомного полиморфизма — строения и числа хромосом. 4)Биохимический подход. В последние десятилетия при изучении природных поп-ций все более широко использ-тся неск-ко подходов, кот-е по применяемым методам можно объединить как биохимические. Часто исследователи ставят знак равенства между изучением биохимического полиморфизма белков и генетикой популяций. 5)Онтогенетичекий подход. Суть онтогенетич-го подхода к изуч-ю поп-ций заключ. в использовании особенностей индивидуал-го развития (т.е.любых особ-тей роста и развития особей) для характ-ки изучаемых поп-ций. 6)Физиологический подход. Физиология изучает любые проявл-я жизнедеят-ти (функционирования) орг-мов. Изучение физиологич-х особенностей особей с целью характеристики поп-ций и составляет содерж-е направления, кот-е может быть названо «популяц-й физиологией». 7)Этологический подход. 2 осн-х направл-я, реализуемых при этологическом исследовании поп-ций: 1) сравнение особенностей поведения особей разных поп-ции и их групп 2) изуч-е индивид-х (внутрипопуляц-х) особенностей поведения.
5.Основные свойства популяции как биологической системы. 1)Поп-ция как череда онтогенезов. Поп-ция — это непрерывный во времени поток онтогенезов связ-ных родством. Это родство может быть либо сетчатым (у размнож-хся половым путем), либо дихотомическим (у аблигатноагамных форм), либо попеременно тем и др. (у форм, половое размнож-е кот-х чередуется с бесполым). Непрерыв-ть поп-ции во времени определяет возможность воспроизвед-я поп-ции в чреде поколений.
2) Целостность поп-ции. Поп-ция всегда целостная система. Она как целое реагирует на разные влияния в эволюцион. масштабе, т. е. на действие любых эволюц-х факторов. Целостность поп-ции опред-тся, в 1-ю очередь, св-вами поп-ции как генетич. системы — реальной связью, подавляющей части ее членов м/ду собой в чреде поколений как более или менее далеких родственников и возмож-ю обмена генетич. инф-цией при скрещивании. Именно связи выступают как главный, объективно цементирующий популяц-ный фактор. 3)Изолированность и многочисленность как основа независимости популяции. Независ-ть поп-ции внутри вида опред-тся, в 1-ю очередь, ее постоянной изоляцией от др. поп-ций. В среднем поток мигрантов в любую поп-цию не превышает в природе немногих %-в на поколение, обычно же много меньше (порядка долей %-та). Это ограничение обмена аллелями между поп-циями и плюс неизбежное несколько различное давление естеств. отбора в каждой поп-ции явл. важными факторами, опред-ми независимость всякой популяции. Независимость поп-ции проявл. и в том, что любая поп-ция вида обладает всеми генами данного вида. Несхожесть между поп-циями касается (с генетич. точки зрения) лишь различий в частотах встречаемости различн. аллелей, но не самих генов. Генофонд во всех поп-циях одного вида одинаков, но аллелофонд- различный.Независ-ть поп-ции определяется и многочисл-тью составляющих ее особей. Только достаточно большая численность определит возмож-ть образов-я и устойчивого существов-я поп-ции как целостной системы, возможность выжив-я этой системы после резких, порой многократных сокращений числен-ти, характ-х для всех живых существ. Достаточн. численность особей, составляющих поп-цию, оказывается одной из важн-х черт поп-ции, определяющей возможность ее длит-ного существов-я в условиях таких колебаний.
4)Структурированность и полиморфность как основа адаптабильности популяции. Поп-ция — не гомогенное образование, а сложноструктурированная система. Любая поп-ция характ-тся специфическойбиохорологической организован-тью. Эта многоплановая структурированность поп-ции определяет одну общую для всех поп-ций и очень важную особенность- полиморфность (в широком смысле). В популяции всегда присутствуют разные по возрасту, полу, морфологии, физиологии и этологии (для животных) группы особей. Не менее 10-20% локусов у каждого индивида оказываются гетерозиготными. Это отражает постоянное присутствие в любой достаточно многочисл-ной группе особей огромного числа различных генетич-х вариантов. При подавляющем большинстве даже необычных воздействий в этих условиях в популяции обычно находятся особи, способные либо выжить, либо процветать. 5) Динамичность поп-ции. Все структуры и границы поп-ции подвижны, динамичны. Непрерывно меняется возрастной состав поп-ции. Исчезнов-е одних генотипов и появл-е др. уже определяют неизбежность постоянного измен-я популяционного аллелофонда. Множество экологич. факторов, постоянно действуя в природе, ежеминутно меняют морфологич-е, физиологич-е, этологич-е и др. характ-ки поп-ции. 6)Уникальность поп-ции. Двух одинаковых поп-ций нет и быть не может как по причине генетич. уникальности особей, их составл-щих, так и по причине невозможности точного повтор-я биотического и абиотического окруж-я в 2-х пространственно раздел-х участках биосферы.
6.Демографические характеристики поп-ции: общая числ-ть. М-ды учёта обилия. Чтобы понять, как функционирует экологич. система, необходимо знать характ-кипоп-ций, составляющих эту систему. Поп-цию в целом характеризуют демографич. характеристики: рождаемость; смертность; структура особей по возрастному составу; численность особей (их обилие).Демографич. характ-ки отражают скорость пр-ссов, происходящих в поп-ции. Знание демографич. характ-тик поп-ции важно для прогнозир-ния возможных изменений, как в самой поп-ции, так и во всем сообществе в целом. Рожд-ть- число особей (яиц, семян и т. д.)—ΔNn, родившихся (отложенных, продуцированных) в поп-ции за некот-й промежуток времени Δt. Рожд-ть явл. важной демографич-й характ-кой. Для поп-ции можно рассчитать 2 показателя рожд-ти: абсолютную и удельную. Абсолютн. рожд-ть - кол-во родившихся особей за к.-либо промежуток времени. Для того чтобы удобнее было сравнивать м/ду собой поп-ции разной числен-ти, величину ΔNn/Δt обычно относят к общему числу особей N в начале промежутка времени Δt. Полученную величину ΔNn/NΔt называют удельной рожд-тью. Удельная рожд-ть - среднее измен-е числ-ти на особь за определ-й интервал времени. Смертность - величина, обратная рожд-ти. Это число погибших в поп-ции особей за ед-цу времени. Подобно рожд-ти, смерт-ть можно выразить числом особей, погибших за данный период или же в виде удельной смерт-ти для всей поп-ции или ее части. Абсолютная (общая) смерт-ть — это число особей, погибших в единицу времени. Удельная смертность выражается отношением абсолютной смерт-ти к числ-ти поп-ции. Возрастная структура поп-ции – соотнош-е в поп-ции особей разных возрастных групп. Календарный возраст – абсолютный возраст, время от момента рождения. Биологический возраст – постадийный возраст, определяющий роль орг-ма в популяц-х пр-ссах.Важной характеристикой при исследовании поп-ции явл. ее возрастная стр-ра, влияющая как на рожд-ть, так и на смерт-ть. Соотношение разных возрастных групп в поп-ции определяет ее способность к размнож-ю и показывает ее перспективу. Числ-ть поп-ции - это общее кол-во особей некот-го вида, присутствующее на той или иной территории. Числ-ть поп-ции - важная экологич. характеристика поп-ции. Число особей в поп-ции имеет огромное эволюционное знач-е. Но важна не общая числ-ть особей в поп-ции, а эффективная числ-ть - репродуктивная числ-ть - та часть поп-ции, кот-я формирует генофонд следующего поколения (генетически эффективная величина).
Методы учёта обилия: 1) М-д прямого (тотального) подсчёт а. Можно применять в случае не только сидячих орг-мов, но и быстро движущихся крупных животных.2) Фотографиров-е. Прямым подсчетом особей на фотоснимках, сделанных с самолета, можно установить размеры поп-ций крупных млекопит-х и морских птиц, собирающихся на открытых пространствах. Можно также использ-ть «фотоловушки», кот-е устанавливают вдоль звериных троп. 3 )М-д изъятия. Этот м-д удобен для оценки числ-ти мелких орг-мов, напр. насекомых, на известной площади или в данном объеме воды. 4) Стандартизированным способом (напр, делая определ-е число взмахов сачком установл-го размера) отлавливают некот-е число животных, подсчитывают их, но не выпускают до конца исследов-я. Процедуру повторяют еще неск-ко раз, при этом с каждым разом число пойманных животных уменьшается. По этим данным строится график, экстраполируя кот-й, получают общую числ-ть животных: она соответствует моменту, когда они перестают попадаться (нулевой ординате), т. е. все особи данного вида теоретически оказываются отловленными и подсчитанными. 5) М-д отлова-выпуска. Этот м-д включ. отлов животного, мечение безвредным для него способом и возвращ-е его на прежнее место в поп-ции. Ч/з некот-е время проводят повторный отлов, при кот-м меченые особи оказываются «разбавленными» теми, что поплись впервые. 6) М-д с использ-ем радиопередатчиков и радиолова. 7)М-д подсчета числ-ти поп-ции, подверженных избирательному промыслу.
7.Плотность поп-ции - число особей, приходящихся на единицу площади или объема. Изменение плотности популяции позволяет сделать вывод о соотношении рожд-ти и смерт-ти, но только в тех условиях, если ареал поп-ции остается неизменным и не происходит ни эмиграции, ни иммиграции особей. Если в кач-ве критерия измен-я численности поп-ции использовать чистую скорость размножения r0, равную среднему числу потомков, производ-х данной особью вида за всю жизнь, то при: r > 1 – поп-ция растущая, r = 1 – поп-ция стабильная, r < 1 – поп-ция сокращающаяся
Колебания числ-ти особей любой поп-ции наз-тся волнами жизни или популяц-ми волнами. Могут быть сезонными (периодическими), то есть обусловл-ми генетически, а также несезонными (апериодическими), то есть обусл-ми непосредственным возд-ем на поп-цию биотических и абиотических факторов. Длина волны жизни прямо пропорц-на продолжит-ти цикла развития орг-ма. Численность поп-ции завис.от множества факторов, кот-е можно условно раздел. на 2 группы: 1)факторы, не зависящие от плотности поп-ции. Напр., абиотические (наиболее важны - климатические). 2)факторы, зависящие от плотности поп-ции: конк-ция, деятельность паразитов, хищников, болезни, пища. Если плотность поп-ции приближается к некот-му верхнему пределу, то вл-е этих факторов усиливается, но в некот-х случаях может и снижаться.

1. Отвечает случаю, когда скорость роста поп-ции уменьшается с увелич-ем ее численности. Это харак-но для большинства поп-ций растений и животных и проявляется 2-мя способами: 1)при увеличении плотности поп-ции - снижение плодовитости; 2)при увеличении плотности поп-ции изменяется возраст наступл-я половой зрелости.
2. Соотв-ет макс-му темпу роста поп-ции при средних, а не при низких значениях плотности. Однако, достигнув макс-го знач-я, скорость роста поп-ции начинает уменьшаться при дальнейшем увеличении плотности поп-ции. Характерно для некот-х птиц, насекомых, видов, для кот-х характерен эффект группы.
3. Наблюд-ся тогда, когда темпы роста поп-ции примерно постоянны при высоких значениях плотности. После достиж-я предельного знач-я плотности поп-ции, темп роста сильно падает. Характ-но для видов с сильными колебаниями численности (мышевидные грызуны, насекомые).
Плотность поп-ции — важный экологич-й показатель пространств-го размещ-я членов поп-ции, а также динамики численности животных, условий изменч-ти и проявл-я естеств. отбора. Плотность поп-ции опред-тся преимущественно степенью благоприятности условий существования вида в данном биотопе и либо важными экологич. факторами окруж. среды, особенно находящимися в минимуме и называется лимитирующими. Поэтому по средней плотности поп-ции можно судить о благоприят-ти местонахожд-е для данного вида. Плотность поп-ции и характер пространственного распредел-я животных закономерно меняются при циклич-х колебаниях числ-ти. Рост плотности поп-ции у большинства видов сопровождается выдел-ем ее сочленами и накоплением во внешней среде продуктов обмена, в т.ч. особых сигнальных в-в, кот-е тормозят или ускоряют рост и развитие, огранич-ют или даже прекращают размнож-е, могут увеличивать подвижность животных и менять их повед-е. В р-тате при высокой плотности поп-ции усиливается рассел-е и может начаться массовая эмиграция. При уменьшении плотности поп-ции эмиграция прекращается, а подвижность неск-ко падает, снова увеличиваясь при чрезмерном разряжении поп-ции, что чревато разрушением внутрипопуляц-х группировок. Одновременно возрастает интенсивность размнож-я.
У каждого вида в завис.от его образа жизни и подвижности (сидячие, оседлые или кочевые, мигрирующие на большие расстояния) существуют оптим-я плотность поп-ции и допустимые пределы ее колебаний, неодинаковые в разных биотопах (макс. и мин. плотность поп-ции). У неподвижных организмов (растения, микроорганизмы, сидячие животные), получающих пищу и кислород из окруж. среды с потоками воды, воздуха, почвенными раств-ми, возможно, а во многих случаях и выгодно примыкание орг-мов др к др. Животные большинства видов держатся поодиночке или небольш. группами (семьями), занимая опред. участки (индивид-е или семейные), кот-е, как правило, примыкают др. к др., иногда частично сочетаясь или перекрываясь.
Плотность поп-ции, соответствующая образу жизни вида и условий его существов-я, поддерживается и регулируется многими механ-ми, эволюционно сложились. Главное знач-е имеет территориальность, т.е. способность осваивать и охранять от вторж-я занятую территорию с помощью активных действий и предупредит-х сигналов (хим-х, визуал-х, акустич-х и др.). Для поддержания групп есть сигналы противополож. значения (привлекающие особей одной семьи или стада).
8.Пространственное распределение популяции. Типы распределения
Пространственная структура — характер распределения особей популяциина занимаемой территории. В природе популяциям свойственны 3 типа пространственного распредел-я особей: случайное, равномерное, групповое. Они формир-ся в завис-ти от степени неоднород-ти среды обитания, биологич. особенностей вида и поведения особей.
Случайное распределение происх-т, если среда обитания относит-но однородна по экологич. условиям. При этом кол-во особей в поп-ции невелико, и биологич-е особенности вида не позволяют им образовывать группы. Напр., у белой планарии, пресновод. полипа гидры, пауков, двустворчатых моллюсков отмечается случайное распредел-е.

Равномерное распред-е наблюд. у видов, жестко конкурир-щих за пищевые ресурсы и территорию. Тенденция к равномерному распределению особей у некоторых животных может быть обусловлена мечением я охраной мест обитания. В природе равномерн. распредел-е встречается довольно редко. Напр., кустарники в пустыне, конкурируя за влагу, распределены довольно равномерно. Поддерживают между собой опред-ю дистанцию некот-е виды хищных рыб, птиц и млекопитающих, охраняющих свои кормовые территории.
Групповое распределение наиб.распространено в природе. Неоднородность среды, огранич-ть местобитания, биологич. особен-ти вида, способы размножения могут приводить к объединению особей в группы. Групповое распределение у растений обусловлено их способами размножения и распространения семян и плодов. Например, некоторые растения образуют крупные, тяжелые плоды (орех лещины, желудь дуба), которые падают рядом с деревом и тут же прорастают, образуя группы. При вегетативном размножении корневищами у растений также формируются группы.
У многих млекопитающих и птиц наблюдается социальное поведение, кот-е приводит к образованию групп с социальной иерархией (стан, става, колонии, табуны, семьи, гаремы). Выживаемость особей в группе повышается благодаря лучшим возможностям для защиты от врагов, обнаружения корма, противост-я неблагоприят. факторам среды, формир-я микроклимата. Напр., стае волков легче охотиться, а табуну лошадей- защищаться от волков. В семьях птиц и млекопитающих благодаря заботе родителей повышается выживаемость потомства. Группа растений способна лучше противостоять ветру, эффективнее использовать воду.
9.Рождаемость. Показатели рожд-ти. Типы рожд-ти. Рожд-ть- число особей (яиц, семян и т. д.)—ΔNn, родившихся (отложенных, продуцированных) в поп-ции за некот-й промежуток времени Δt. Рожд-ть явл. важной демографич-й характ-кой. Для поп-ции можно рассчитать 2 показателя рожд-ти: абсолютную и удельную.Абсолютн. рожд-ть - кол-во родившихся особей за к.-либо промежуток времени. Для того чтобы удобнее было сравнивать м/ду собой поп-ции разной числен-ти, величину ΔNn/Δt обычно относят к общему числу особей N в начале промежутка времени Δt. Полученную величину ΔNn/NΔt называют удельной рожд-тью. Удельная рожд-ть - среднее измен-е числ-ти на особь за определ-й интервал времени. Поскольку в течение исслед-го промежутка Δt величина рожд-ти может меняться, этот промежуток стараются сделать по возможности короче. Переходя на язык математики, можно записать, что при Δt→0 выражение ΔNn/NΔt примет вид aNn/Ndt = b. Получ-ю величину b наз-т также мгновенной удельной рожд-тью. Размерность ее— «единица времени-1». Единица времени, выбранная для оценки рожд-ти в той или иной поп-ции, изменяется в завис-ти от интенсивности размнож-я исследуемых орг-мов. Для растущей в оптимальных лаб. условиях поп-ции бактерий такой единицей может быть час, для поп-ции планктонных водорослей — сутки, для многих насекомых— неделя или месяц, а для крупных млекопит-х—год. Рожд-ть по определению может быть величиной положит-й или +0. Величина рожд-ти завис.от многих причин. Большое знач-е имеет доля особей, способных в данный период к размнож-ю, что опред-тся соотношением полов и возрастных групп. Важна также частота последовательности генераций. Так, среди насекомых различают моновольтинные и поливольтинные виды. 1-е дают 1, 2-е – неск-ко генераций за год. По числу периодов размножения в течение жизни различ. моноциклические и полициклические виды. Моноцикличность, или однократное размнож-е, свойственна обычно видам с короткой продолжит-тью жизни в половозрелом состоянии (лососевые рыбы, поденки, майские жуки и др. насекомые). Полицикличность характ-тся повторным размнож-ем особей и присуща большинству позвоночных живот-х и ряду беспозвоночных, напр. ракообразным. Экологи выд. макс. рожд-ть в условиях отсутствия лимитирующих экологич-х факторов (добиться этого невозможно). Макс. рожд-ть- теоретически возможный максимум скорости образов-я новых особей в идеальных условиях. Размнож-е орг-мов сдерживается только их физиологическими особенностями. Макс. рожд-ть — понятие теоретич-е. Ни 1 вид в природе не может бесконтрольно и безгранично размножаться. В отличие от макс. экологическая, или реализованная, рожд-ть характ-ет прирост или увеличение числ-ти поп-ции при фактических или специфич-х условиях среды. Любая поп-ция теоретически способна к неогранич. росту числ-ти, если ее не лимитируют факторы внеш. среды. В таком гипотетическом случае скорость роста поп-ции будет зависеть только от величины биотического потенциала, свойственного виду.
10.Плодовитость, её отличие от фертильности. Плодовитость — способность к размнож-ю. В биологии и демографии это возможная репродуктивная способность орг-зма или поп-ции, измеряемая числом гамет (яиц), семян, спор или отростков растений. Плодов-ть опред-тся наследственностью и средой и явл. осн. мерой приспособленности особи.
Д. Лэ к установил на примере птиц, что у них отбор благоприятствует не макс. Плодов-ти, а наиболее эффективной, т. е. тому колич-ву яиц, при кот-м потомство оказывается наиболее жизнеспособным. Большая плод-ть вырабатывается у видов также в условиях более высокой смерт-ти, особенно при сильном давлении со стороны хищников. Отбор на плодов-ть компенсирует высокую норму гибели в поп-циях. Поэтому при высокой плод-ти рост численности поп-ции может быть весьма низким. Вразличных поп-циях 1-го вида плод-сть обычно тем выше, чем более неблагоприятны занимаемые ими места обитания.
Плодовитость животных непосредственным образом связана с размерами самок, продуцирующих яйца. В толерантных условиях абсолютная плодовитость у пойкилотермных животных конкретных видов возрастает с увеличением размеров их тел. Степенная зависимость плодовитости от массы животных справедлива в пределах лишь определенного размерно-возрастного диапазона. Нижняя граница этого диапазона определяется размерами (массой), которых животные достигают ко времени наступления половозрелости или первого вымета молоди у живородящих гермафродитов. По достижении самок животных определенного размера или возраста число потомков у них начинает уменьшаться, несмотря на продолжающееся увеличение размеров или массы. Это связано с ослаблением репродуктивной способности у особей старших возрастов.
Коэффициент общей плодовитости –демографич-й показатель: отнош-е числа родившихся за год к общей численности женщин в возрасте от 15 до 49 лет, выраженное в промилле (‰): относится к группе специальных показателей рождаемости.
Плод-ть меняется с возрастом, закономерно колеблется по сезонам (у видов с повторным размнож-ем) и в разные годы в завис-ти от степени обеспеченности пищей, плотности поп-ций, климатич-х и др. факторов. Является предметом изучения в популяционной экологии.
У разных групп животных плод-ть очень различна, особенно у видов с разными способами размнож-я. Некот-е насекомые откладывают до неск-х сотен тысяч и даже миллионов яиц. Животные с большой продолжит-тью жизни и высокой степенью заботы о потомстве производят гораздо меньше гамет и соответственно потомства.
Плодов-ть это только возможность произвести потомство, зависящая от продукции гамет, оплодотворения и вынашивания до рождения, и поэтому отличается от фертильности, мерой кот-й явл. реальное кол-во потомков, напр. скорость рождения в поп-ции выражаемая числом новорожденных на 1000 особей в год.
Ферти́льность — способность половозрелого орг-ма производить жизнеспособное потомство. Противоположно понятию «стерильность». В физиологич. смысле под фертильностью понимают способность женск. или мужск. орг-ма к участию в оплодотворении.Чаще всего отсутствие потомства у супружеской пары обусловлено бесплодием одного из супругов.
11.Смертность. Типы смертности. Смертность - величина, обратная рожд-ти. Это число погибших в поп-ции особей за ед-цу времени. Подобно рожд-ти, смерт-ть можно выразить числом особей, погибших за данный период или же в виде удельной смерт-ти для всей поп-ции или ее части.
Абсолютная (общая) смерт-ть — это число особей, погибших в единицу времени (?Nm).
Удельная смертность (d) выражается отношением абсолютной смерт-ти к числ-ти поп-ции:
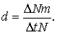
Абсолютная и удельная смерт-ть характеризуют скорость убывания числ-ти поп-ции вследствие гибели особей от хищников, болезней, старости и т. д.
Для анализа роста числ-ти поп-ции обычно пользуются данными, отражающими величину смерт-ти. Для этого сост-т так называемые таблицы и кривые выживаемости - определяют, как смерт-ть распределяется по возрастам. В осн. смерт-ть и возрастная стр-ра видов завис.от того, какие шансы для выжив-я имеют особи в различ. возрастных группах. Наглядн. Инф-цию дает кривая выжив-ти отдел. видов, полученная путем подсчета %-та живых особей в отдельной поп-ции. Для большинства видов такая кривая обычно может быть1из 3-х наиб.типичных вариантов.

Рис. Различные типы кривых выживаемости
У видов, для кот-х роль внеш. факторов в смерт-ти невелика, кривая выжив-я характ-ся небольшим пониж-ем до опред. возраста, после кот-го происх. резкое падение вследствие естеств-й (физиологич-й) смерт-ти. Кривая I типа довольно выпуклая и соответствует ситуации, при кот-й смерт-ть ничтожно мала в течение большей части жизни, но затем резко возрастает, и за короткий срок наблюд-ся гибель всех особей (тип дрозофилы: именно его демонстрируют дрозофилы в лаб. условиях). Кривая II типа предст. собой диагональ и соответ-т постоянной, т.е. независ-й от возраста смер-ти в течение всей жизни (рыбы, пресмыкающииеся, птицы, многолетние травянистые растения). Этот тип принято называть типом гидры - для нее свойственна кривая выжив-я, приближающаяся к прямой линии.
Для многих видов характ-на высокая смерт-ть на ранних стадиях онтогенеза. Кривая III типа сильновогнутая и отражает массовую гибель особей в начальн. период жизни, а затем относит-но низкую смерт-ть оставшихся орг-мов. У таких видов кривая выжив-я характ-тся резким падением в области младших возрастов. Данный тип кривой наз-ют типом устрицы, потому что у этого моллюска на 1 из стадий развития имеется планктонная личинка, и на этой стадии наблюд. чрезвычайно высокая смерт-ть особей. Свойственна донным беспозвон-м, имеющим планктонных личинок, многим орг-мам, характер-мся большой плодовитостью и отсутствием заботы о потомстве. Реально встречающиеся кривые выживания нередко представляют собой некоторую комбинацию указанных выше типов
12.Продолжительность жизни. Для каждого вида орг-мов существует некот-я макс. возможная продолжительность жизни. В природе до возраста, близкого к макс-му, доживает ничтожно малая доля особей, и поэтому при анализе динамики природных поп-ций величина макс-й продолжительности жизни представляет интерес только как крайняя точка отсчета при построении кривых и таблиц выжив-я. По существующим оценкам максимальная продолжит-сть жизни варьируется очень широко: от десятков минут у бактерий, находящихся в оптим-х лаборат-х условиях, до неск-х тысячелетий у некот-х видов древесных растении. Обычно продолжит-сть жизни крупных животных и растений существенно больше, чем мелких, но из этого правила в пределах самых разных групп орг-мов встречаются и исключ-я. Рукокрылые в сравнении с др. мелкими млекопитающими могут жить довольно долго.Продолжит-сть жизни орг-мов 1 вида может сильно изменяться в завис-ти от условий существов-я. Так, напр., пойкилотермные животные, как правило, живут дольше при низк. температуре, чем при высокой (конечно, в том случае, если темп-ры эти не выходят за пределы толерантности). Для некот-х беспозвоночных, напр. ветвистоусых рачков дафний, известна завис-ть макс. продолжит-сти жизни от условий питания: при поддержании в эксперименте постоянной высокой концентрации пищи продолжит-сть жизни дафнии меньше, чем при умеренном кормлении, но больше, чем в условиях крайне низкой концентрации пищи, когда животные испытывают сильное голодание. Причины такого сниж-я не очень понятны, но не исключено, что оно связ. с усилением ф-ции размнож-я: укорачивание жизни есть «плата» за более интенсивное размнож-е.
Средняя продолжит-сть жизни в сравнении с макс-й показатель более информативный для эколога, но и эта величина сама по себе не может заменить сведений о характере распредел-я смертности по возрастам, сведений, кот-е обычно представляют в форме таблиц выживания. Попытка сост-ть таблицы была предпринята Джоном Грантом. Исходн. материалом для расчетов Гранта служили данные по смертности жителей Лондона. 1-я таблица выжив-я, похожая на современные, была сост-на в 1693 г. Э. Галлеем для г.Бреслау, численность населения в кот-ом сохранялась стационарной. В ряде случаев самый простой способ постр-я таблиц выжив-я — это подробно