Весьма замечательно, что сложные схемы связей, обеспечивающих избирательность по отношению к ориентации и направлению движения, а также другие особые свойства, должны быть представлены в виде двух копий. И, пожалуй, еще более поразительно, что все это можно наблюдать у новорожденных животных. Данные схемы связей в основном врожденные и, по-видимому, детерминированы генетически. (Однако в одном специальном отношении некоторое дозревание бинокулярных связей осуществляется главным образом после рождения.)
Теперь мы обратимся к рассмотрению закономерностей группировки клеток в коре. Каким образом распределены там клетки со сходными характеристиками - близкие по степени сложности, локализации рецептивного поля и доминированию глаз - сгруппированы вместе или разбросаны в случайном порядке? Из приводимого описания будет очевидно, что сходные по сложности клетки имеют тенденцию группироваться в слои, причем клетки с радиальной симметрией лежат внизу слоя IV, простые клетки - прямо над ними, а сложные клетки - в слоях II, III, V и VI. Сложные клетки можно подразделить еще на несколько категорий, и те, которые находятся в одном слое, в ряде отношений сильно отличаются от клеток других слоев.
Эти различия между слоями приобретают особый интерес в аспекте следующего важного открытия, подтвержденного рядом физиологов и анатомов в последние несколько десятилетий: волокна, выходящие из разных слоев коры, имеют различные места назначения. Так, самый глубокий слой зрительной коры - слой VI - проецируется главным образом (а возможно, и исключительно) обратно в латеральное коленчатое тело; слой V проецируется в верхнее двухолмие, зрительный отдел среднего мозга; слои II и III направляют свои волокна в другие части коры. Такая определенность в локализации проекций каждого слоя, возможно, заслуживает того, чтобы рассматривать ее как третье важное прозрение относительно организации коры.

Ориентационные колонки удалось выявить в виде уловимых анатомических структур с помощью дезоксиглюкозных радиоавтографов, полученных авторами и М. Страйкером. Обезьяне вводили дезоксиглюкозу с радиоактивной меткой; она поглощалась преимущественно работающими клетками, и в них накапливались продукты первых этапов метаболизма. Сразу после инъекции животному предъявляли решетку из вертикальных полос, в связи с чем клетки, реагирующие предпочтительно на вертикальные линии, были самыми активными, а затем стали самыми радиоактивными. На этом срезе перпендикулярные к поверхности области активных клеток представляют собой узкие полосы, расположенные на расстоянии около 0,5 мм. Слой IV (не проявляющий избирательности в отношении ориентации), как и ожидалось, однороден по радиоактивности.
Следующий варьирующий параметр стимуляции, который нужно рассмотреть, - это положение рецептивного поля в поле зрения. Описывая латеральное коленчатое тело, мы указывали, что в каждом его слое образуется упорядоченная топографическая карта контралатеральной половины поля зрения. В проекции латерального коленчатого тела на зрительную кору этот порядок сохраняется, в результате чего создается кортикальная карта поля зрения. При наличии такой упорядоченной карты не удивительно, что соседние клетки в этой части коры всегда имеют рецептивные поля, располагающиеся в непосредственной близости; в действительности, они обычно перекрываются. Если в кору под прямым углом к поверхности погружать микроэлектрод и регистрировать активность клетки за клеткой, сделав до 100 или 200 отведений во все более глубоких слоях, то опять же рецептивные поля будут большей частью перекрываться, причем каждое новое поле будет накладываться на все предыдущие. Область, занимаемая всей «пачкой» полей, обычно в несколько раз превышает размер одного типичного поля.
Размеры этих рецептивных полей несколько варьируют. Некоторые вариации соотносятся со слоистостью: наибольшие поля при любом погружении имеют тенденцию обнаруживаться в слоях III, V и VI. Однако наиболее важные вариации коррелируют с эксцентриситетом, или расстоянием рецептивного поля клетки от центра взора. Размеры полей и величина соответствующего разброса в тех областях коры, где картирована область центра взора, выглядят крошечными по сравнению с размерами и разбросом в тех частях, где картирована далекая периферия. Пачку накладывающихся полей, которые картируются при погружении, начинающемся в любой точке коры, мы называем «агрегатным» полем этой точки. Очевидно, что величина агрегатного поля является функцией расстояния от центра взора.

Картина распределения предпочтительных ориентации, если ее рассматривать в фас, неожиданно оказывается весьма сложной. Этот радиоавтограф получен с помощью дезоксиглюкозы со среза, тангенциального к несколько изогнутым слоям коры. Более темные области соответствуют однородно радиоактивному слою IV. В других слоях ориентационные области выглядят как сложным образом изогнутые полосы, напоминающие стены рассматриваемого сверху лабиринта, однако расстояния между полосами примерно одинаковы.
Если электрод входит наклонно, почти параллельно поверхности, снова выявляется разброс в позиции полей от точки к точке, но теперь этот разброс накладывается на систематическое смещение позиции поля, направление которого диктуется топографической картой поля зрения. При этом обнаруживается интересная закономерность: оказывается, что продвижение электрода примерно на 1-2 миллиметра всегда приводит к такому смещению в поле зрения, которого достаточно для перехода в совершенно новую область. Короче, величина продвижения в поле зрения примерно соответствует при этом размеру агрегатного рецептивного поля. Для первичной зрительной коры это справедливо, где бы ни производилось погружение. В центре взора поля и их разброс малы, но таковы же и смещения, соответствующие продвижению на миллиметр вдоль коры. С увеличением эксцентриситета (по мере приближения к границам поля зрения) как поля и их разброс, так и смещения, становятся пропорционально больше. Получается, что повсюду кусочек коры протяженностью в один или два миллиметра является тем блоком, который обслуживает область поля зрения, эквивалентную величине агрегатного поля.
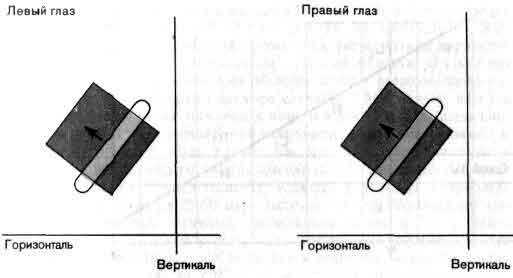
Бинокулярные клетки могут возбуждаться как каждым глазом независимо, так и более сильно - двумя глазами вместе. Здесь представлены относящиеся к левому и правому глазам рецептивные поля сложной клетки, контролирующей область в верхнем левом квадранте поля зрения. (Оси координат соответствуют горизонтальному и вертикальному меридианам поля, пересекающимся в точке фиксации.) Два данных рецептивных поля идентичны, но сила реакции может зависеть от того, какой глаз стимулируется: правый или левый. Глаз, оказывающий более сильное влияние, называется доминантным.
Из этого наблюдения следует предположение о способе, с помощью которого кора решает такую фундаментальную проблему: как сделать, чтобы зрительная картина анализировалась детально в центральной части и намного грубее на периферии. В сетчатке, перед которой стоит такая же проблема, по очевидным оптическим причинам число миллиметров, соответствующих градусу поля зрения, постоянно. Сетчатка обрабатывает центральные участки более детально благодаря тому, что имеет огромное количество ганглиозных клеток, каждая из которых обслуживает крошечную область в центре поля зрения; слой ганглиозных клеток в центральной части сетчатки относительно толст, тогда как в периферических частях сетчатки он очень тонок. В то же время, было, по-видимому, желательно, чтобы кора везде имела одинаковую толщину. Здесь нет никаких оптических ограничений типа накладываемых на сетчатку, и потому площади просто распределяются в соответствии с проблемами, которые должны решаться.
На каждом квадратном миллиметре коры предположительно действуют примерно такие же механизмы, как на любом другом. Несколько тысяч волокон из коленчатого тела входит в такую функциональную ячейку коры, и что-то около 50000 волокон выходит из нее, независимо от того, представлена ли здесь малая часть зрительного мира очень детально или большая по размеру часть соответственно менее детально. Как мы указывали вначале, предположение об однородности коры возникает при взгляде на окрашенные срезы. Оно убедительно подтверждается, когда мы исследуем архитектонику дальше, специально обращая внимание на чувствительность к ориентации или на доминирование глаз.

В физиологических исследованиях была выявлена группировка клеток в соответствии с доминированием глаз. При некотором произвольном погружении в кору под прямым углом к поверхности (1) микроэлектрод может встретить только клетки, которые оказывают предпочтение левому глазу (Lr), а в слое IV - клетки, которые возбуждаются только левым глазом (L); при другом вертикальном погружении (2) для всех клеток доминантным будет правый глаз (R,), а в слое IV клетки будут возбуждаться исключительно правым глазом (R). При наклонном погружении будет наблюдаться регулярное чередование доминирования глаз. Результаты многократных погружений наводят на мысль, что кора подразделена на области с поперечным сечением шириной около 0,4 мм и со стенками, перпендикулярными поверхности и слоям коры - колонки глазодоминантности.
Относительно ориентации мы исследовали группировки клеток точно так же, как это мы делали при рассмотрении локализации полей, т. е. сначала изучали пары клеток, расположенных в непосредственной близости друг от друга. Две такие клетки почти всегда имеют одинаковые оптимальные ориентации стимулов. Если электрод вводится в направлении, перпендикулярном к поверхности, все клетки вдоль пути его проникновения имеют одинаковые или почти одинаковые предпочтительные ориентации (кроме клеток в глубине слоя IV, которые совсем не имеют оптимальных ориентации). Однако при двух погружениях перпендикулярно поверхности, произведенных на расстоянии в один миллиметр или около того, обнаруженные две ориентации, как правило, различны. Следовательно, кора должна подразделяться на какие-то вертикальные ячейки, внутри которых оптимальная ориентация одинакова для рецептивных полей всех клеток. Когда мы натолкнулись на эту систему почти 20 лет назад, она заинтриговала нас, поскольку она так хорошо соответствовала иерархическим схемам, предложенным нами для объяснения того, каким образом сложные клетки снабжаются входными сигналами от простых: эти схемы включали связи с клетками, поля которых покрывают одну и ту же часть поля зрения и которые реагируют на одну и ту же ориентацию линий. Казалось совершенно разумным, что прочно взаимосвязанные клетки должны быть сгруппированы вместе.
Если кора разбита на области с постоянной для рецептивных полей оптимальной ориентацией, можно ли сказать что-нибудь большее о трехмерной форме этих областей, кроме того, что их стенки перпендикулярны поверхности? Соотносятся ли соседние области сколько-нибудь систематическим образом, или обслуживающие всевозможные направления колонки разбросаны по коре случайно? Мы начали изучать ни вопросы, просто вводя электрод в кору наклонно или параллельно поверхности. Когда мы первый раз поставили такой эксперимент в 1961 г., результат был столь удивителен, что мы с трудом в него поверили. Вместо случайного набора сменяющихся ориентации наблюдалась изумительная упорядоченность. Каждый раз, когда микроэлектрод продвигался всего на 25-50 мкм (тысячных долей миллиметра), оптимальная ориентация менялась небольшим скачком, в среднем примерно на 10°; скачки в одном направлении - по часовой стрелке или против нее - продолжали наблюдаться в довольно большом диапазоне углов, что-то от 90 до 270°.
С тех пор как было сделано это первое наблюдение, мы находим аналогичную упорядоченность почти у каждой обезьяны. Либо стабильно регистрируется поворот ориентации, либо, реже, встречаются участки, в пределах которых ориентация остается постоянной. Последовательные изменения ориентации достаточно малы, поэтому трудно быть уверенным в том, что области постоянной ориентации имеют конечные размеры; возможно, что по мере продвижения электрода вдоль коры оптимальные направления меняются в каком-то смысле непрерывно.
У нас все больше нарастал интерес к трехмерной форме этих подобластей. Уже из рассмотрения одной геометрии очевидна возможность существования малых или нулевых изменений в любом направлении при горизонтальном или тангенциальном проникновении к параллельным слоям ткани, содержащим клетки со сходной специфичностью, и таким, что каждый слой перпендикулярен поверхности. Слои не обязательно должны быть плоскими как ломти хлеба; при взгляде сверху некоторые из них имеют вид завихрений, с помощью которых легко объяснить инверсии в направлении поворота ориентации. Запись от множества клеток по ходу нескольких параллельных погружений электрода, по-видимому, подтверждает такое предположение, однако с помощью микроэлектрода мы могли исследовать не более чем крошечный участок мозга.
К счастью, как раз тогда, когда нам было нужно, был изобретен идеальный анатомический метод. Это был метод оценки активности мозга с помощью 2-дезоксиглюкозы, предложенный Л. Соколовым (L. Sokoloff) и его группой из Национального института охраны психического здоровья (метод этот описан в данном выпуске в статье Л. Ивер-сена). В основу метода положен тот факт, что клетки мозга используют в качестве источника метаболической энергии главным образом глюкозу и что близкородственное ей соединение, 2-дезоксиглюкоза, может в какой-то степени ее имитировать. Если в организм животного ввести дезоксиглюкозу, она будет так же интенсивно поглощаться нейронами, как и сама глюкоза, и чем выше активность нейрона, тем интенсивнее будет поглощение. Соединение начинает включаться в метаболический цикл, но по причинам, которые лучше известны биохимикам, процесс останавливается на метаболитах, не способных проникать через клеточные мембраны и потому накапливающихся в клетках.

Анатомическое подтверждение существования колонок глазодоминантности было получено с помощью различных методов окраски и методом аксонного транспорта с применением радиоавтографии. Данный составной радиоавтограф, проявляющий узор на площади шириной около 10 мм, был получен путем изготовления и совмещения срезов из областей, представляющих слой IV в некотором числе параллельных сечений.
Метод Соколова состоит в том, что животному вводят меченную радиоактивным изотопом (14С) дезоксиглюкозу, затем воздействуют раздражителем, предназначенным активировать определенные нейроны, и сразу же определяют радиоактивность ткани мозга; в результате обнаружатся активные области, где клетки поглотили больше дезоксиклюкозы, чем в областях, остававшихся в покое. Обычный способ исследования мозга для этой цели - изготовить тонкие срезы (как это делается для гистологического исследования) и прижать эти срезы к фотопластинке, чувствительной к радиактивному излучению. После проявления все участки, находившиеся в контакте с радиоактивным материалом, будут видны как темные массы осажденных зерен серебра. Вместе с М. Страйкером (М. Stryker) мы приспособили метод Соколова к нашим проблемам, вводя анестезированному животному дезоксиглюкозу и затем двигая изображения черно-белых вертикальных полос туда и обратно на расстоянии 1,5 м от животного в течение 45 минут. Затем мы готовили срезы мозга либо перпендикулярно поверхности коры, либо параллельно ей.
Реконструкция узора глазодоминантности по всей наружной части правой первичной зрительной коры была выполнена авторами и С. ЛеВэем по сериям срезов, окрашенных методом серебрения, разработанным ЛеВэем. Левая граница проходит по медиальному краю затылочной доли, где кора загибается внутрь.
Полученные радиоавтографы быстро подтвердили физиологические результаты. На срезах, перпендикулярных поверхности, выявились узкие полосы радиоактивности, встречающиеся примерно через каждые 570 мкм (грубо говоря, полмиллиметра) и простирающиеся по всей толще коры. Очевидно, это и были области, которые содержали клетки, реагирующие на вертикальные линии. Глубокая часть слоя IV была однородно радиоактивна, как мы и ожидали на основании того факта, что клетки в этом слое имеют рецептивные поля с радиальной симметрией и не проявляют избирательности по отношению к ориентации.
На срезах, параллельных поверхности, выявилась неожиданно сложная система чередующихся полос, часто образующих завихрения, неоднократно расходящихся и воссоединяющихся и только местами образующих регулярные параллельные переслаивания. Что тут было особенно удивительно, так это равенство расстояний между полосами по всей коре. Это прекрасно согласовалось с идеей об однородности коры. Кроме того, величина расстояний между полосами хорошо увязывалась с идеей о том, что кортикальные механизмы повторяются, по крайней мере через каждый миллиметр. Если бы расстояние, требующееся для поворота от вертикали через 180° назад к вертикали, было равно, скажем, 10 миллиметрам, значительная часть коры не содержала бы клеток, чувствительных к какой-то заданной ориентации, что вело бы к фрагментарному и чрезвычайно причудливому представлению зрительной картины.
Последняя переменная, вклад которой в архитектонику надо рассмотреть, - это предпочтение того или другого глаза. При микроэлектродных исследованиях почти неизменно оказывалось, что соседние клетки «предпочитают» один и тот же глаз. Если при вертикальном погружении первая клетка, на которую мы натыкались, предпочитала правый глаз, то также было и со всеми другими клетками вплоть до основания слоя VI; если же первая клетка предпочитала левый глаз, то же было верно и для остальных. Вероятности предпочтения того или другого глаза при произвольном отведении были равны. (Поскольку клетки слоя IV монокулярны, тут нужно говорить не о предпочтении, а о монополии глаз.) Если электрод вводили наклонно или горизонтально, области предпочтения левого и правого глаз чередовались, причем переключения происходили довольно резко - примерно через каждые полмиллиметра. Таким образом, в коре обнаружилась вторая система подобластей с вертикальными стенками, простирающимися через всю ее толщу. Система глазодоминантности, по-видимому, совершенно независима от ориентационной системы, так как при наклонном или тангенциальном введении электрода две последовательности не имеют очевидного отношения друг к другу.
Происхождение колонок глазодоминантности, как их потом назвали, представляется весьма простым. Окончания волокон коленчатого тела, часть которых обслуживает левый глаз, а часть - правый, группируются на входе в кору таким образом, что в слое IV перемешивания нет. Это приводит к появлению «лево»- и «правоглазых» участков, располагающихся с интервалами около полмиллиметра. Нейрон, находящийся ниже или выше слоя IV, образует связи в этом слое в радиусе примерно до миллиметра по всем направлениям. По-видимому, самые прочные связи образуются с областью слоя IV, наиболее близкой к данному нейрону, так что, по-видимому, на него будет оказывать доминирующее влияние тот глаз, который «обслуживает» эту область.
И снова нам было очень интересно выяснить, как эти «левоглазые» и «правоглазые» области могут выглядеть в трех измерениях. То чередование, которое обнаруживалось физиологически, можно было объяснить и геометрически. Ответ сначала последовал из работ, выполненных методом дегенерации с применением серебрения, предложенным для картирования связей У. Наута (W. Nauta) из Массачусетского технологического института. Потом мы нашли три других независимых анатомических метода для демонстрации этих колонок.

Участок коры площадью около одного квадратного миллиметра и высотой в два миллиметра (окрашенный столбик) можно рассматривать как элементарную ячейку первичной зрительной коры. Она содержит один набор ориентационных колонок, обслуживающих все направления, и один набор слоев глазодоминантности, обслуживающих оба глаза. Эта структура многократно повторяется по всей первичной зрительной зоне. Границы можно проводить произвольно (по правому или по левому глазу, по вертикальной, горизонтальной или наклонной ориентации); представление данных подслоев в виде плоских пластин, пересекающихся под прямыми углами, является крайним упрощением.
Один из особенно эффективных методов (поскольку он позволяет наблюдать на одном животном распределение колонок по всей первичной зрительной коре) основан на явлении аксонного транспорта. Процедура сводится к инъекции в нервную ткань меченой аминокислоты. Аминокислота поглощается телом клетки, по-видимому, включается в белок и затем транспортируется поаксону к его окончаниям. Когда мы инъецировали аминокислоту в один глаз обезьяны, ганглиозные клетки сетчатки поглощали ее и транспортировали по своим аксонам — волокнам зрительного нерва. После этого мы исследовали места назначения этих волокон в латеральном коленчатом теле, покрывая срезы тканей серебряной эмульсией и проявляя ее. Оказалось, что радиоактивная метка отчетливо выделяется в трех взаимно дополняющих друг друга слоях коленчатых тел.
Однако этот метод в обычном применении не дает возможности проследить путь от терминали одного аксона через синапс в следующий нейрон и к его окончаниям, а мы хотели проследить весь путь через кору. В 1971 г. Б. Графштейн (В. Grafstein) из Медицинского колледжа Корнеллского университета показала, что после инъекции достаточно большого количества радиоактивного материала в глаз мыши часть его выходит из терминалей зрительного нерва, поглощается клетками коленчатого тела и транспортируется по их аксонам в кору. Мы подумали, что сходная инъекция в комбинации с методом радиоавтографии позволит выявить в IV слое зрительной коры окончания волокон клеток коленчатого тела, принадлежащие одному глазу.
Наша первая попытка закончилась весьма плачевно: в слое IV были видны лишь слабые тени из нескольких зерен серебра. Только после нескольких недель мы осознали, что обратившись к наблюдению под микроскопом в условиях темного поля, можно извлечь выгоду из свойства зерен серебра рассеивать свет, благодаря чему чувствительность метода возрастет. Мы позаимствовали темнопольный конденсор, и когда взглянули на наш первый слайд в микроскоп, там в слое IV во всей своей красе сияли периодические структуры, выявленные метками (см. верхний рисунок на стр. 166).
Следующим нашим шагом была попытка увидеть картину, так сказать, «в лицо», делая срезы коры параллельно поверхности. Кора обезьяны куполообразна, так что на срезе, параллельном поверхности и тангенциальном слою IV, этот слой выглядит как круг или овал, а на срезе, сделанном ниже слоя IV, его сечение представлено в виде кольца. Монтируя вместе серии таких овалов и колец из набора срезов, можно реконструировать картину на большой площади коры.
Из такой реконструкции сразу стало очевидно, что общий план организации - это чередование параллельных полос, которые представляют окончания, принадлежащие глазу, подвергшемуся инъекции, и промежутков, которые представляют другой глаз. Полосы не так регулярны, как на обоях. (Время от времени мы напоминали себе, что это все-таки биология!) Тут и там полоса, представляющая один глаз, разветвляется на две полосы или кончается тупиком в точке, где ветвится полоса другого глаза. Нерегулярности наиболее обычны вблизи центра взора и вдоль линии, представляющей горизонт. Полосы по всей видимости всегда перпендикулярны к границе между первичной зрительной корой и ее соседом - полем 18, и здесь регулярность наибольшая. Такое общее правило, по-видимому, применимо к мозгу всех макаков, хотя узоры варьируют от одного индивидуума к другому и даже у одной и той же обезьяны от одного полушария к другому.

Гипотетическая картина корковой активности, которая может быть результатом стимуляции левого глаза одним коротким отрезком горизонтальной линии, помешенным в верхнем левом квадранте поля зрения, показана цветными штрихами на схеме участка правой коры, рассматриваемой в фас. Область, в которую поступают входные сигналы от объекта в поле зрения, обведена пунктирной черной линией. Если колонки глазодоминантности и ориентационные колонки расположены так, как это здесь изображено, из всех клеток активируются те, которые оптимально реагируют на приблизительно горизонтальные стимулы, предъявляемые левому глазу.
Ширина комплекта из двух полос постоянна, около 0,8 мм, по всей первичной зрительной коре, что еще раз подчеркивает однородность коры. Опять же эта ширина прекрасно согласуется с той идеей, что в пределах одного квадратного миллиметра коры должны содержаться все механизмы, необходимые для «присмотра» за областью, размером с агрегатное поле. Два описанных выше метода — применение меченой дезоксиглюкозы и транспорт аминокислоты, имеют огромное достоинство в том отношении, что они взаимно совместимы, в связи с чем мы можем применять их одновременно, один — для картирования ориентационных полос, а другой — для выявления колонок глазодоминантности. Число препаратов мозга, исследованных к настоящему времени, слишком мало, для того чтобы можно было сделать те или иные окончательные выводы, но пока эти две системы полос кажутся совершенно независимыми; они и не параллельны, и не перпендикулярны, а пересекаются случайным образом.
Функция, выполняемая колонками глазодоминантности, пока остается тайной. Мы знаем, что нейроны со всеми градациями предпочтения глаза имеются по всей бинокулярной части поля зрения, и возможно, что некоторая регулярная упорядоченная система конвергенции входов гарантирует однородность распределения, благодаря чему ни один глаз не окажется случайно выделенным ни в одном месте. Зачем нужны повсеместно все эти градации предпочтения глаза, само по себе неясно; мы можем только догадываться, что это имеет какое-то отношение к восприятию глубины.
Если собрать вместе все, что стало известно о первичной зрительной коре, будет ясно, что элементарным участком коры нужно считать блок площадью примерно в квадратный миллиметр и глубиной два миллиметра. Знать организацию такого кусочка ткани — это значит знать организацию всего поля 17; целое должно быть в значительной степени простым повторением этой элементарной единицы. Конечно, данную элементарную единицу не следует рассматривать как отдельный изолированный блок. С чего начинать отсчет ориентационных колонок — с колонки, представляющей вертикальную ориентацию, или наклонную или горизонтальную, - совершенно безразлично; точно так же все равно, с какой пары начинать последовательность полос глазодоминантности: левый глаз - правый глаз или правый глаз - левый глаз. Это же справедливо для любой ячейки кристалла хлористого натрия и для любого сложного повторяющегося узора типа тех, какие печатают на обоях.
На что же тогда становится похожей зрительная картина, когда она проецируется на зрительную кору? Предположим, что животное фиксирует свой взгляд на некоторой точке и что единственный объект в поле зрения - прямая линия выше и чуть левее той точки, к которой прикован взгляд. Если бы каждая активная клетка должна была загораться и если бы мы могли стоять над корой и смотреть на нее сверху, какую картину мы бы увидели? Чтобы сделать задачу более интересной, предположим, что картина рассматривается только одним глазом. Ввиду только что описанной архитектоники объект предстанет не в виде линии, а всего лишь в виде набора регулярно расположенных обрывков. Это рассуждение можно проверить непосредственно, если предъявлять обезьяне, у которой один глаз закрыт, набор вертикальных полос, а затем, применив дезоксиглюкозу, изготовить радиоавтограф. Полученная картина не будет большим сюрпризом: это будет набор регулярно расположенных пятен, который будет отражать пересечение двух систем колонок. Вообразите удивление и замешательство маленького гномика, если бы он увидел такую версию внешнего мира!

Реальный образец активности коры, которая была вызвана предъявлением одному левому глазу решетки из вертикальных полос. Полученный с помощью дезоксиглюкозы радиоавтограф тангенциального среза верхних слоев коры. Картина регулярно расположенных темных участков радиоактивности отражает пересечение систем колонок глазодоминантности и предпочтительной ориентации.
Почему эволюция пришла к тому, чтобы взять на себя труд изобрести столь сложную схему, - это вопрос, который продолжает волновать нас. Возможно, самое подходящее объяснение состоит в том, что системы колонок — это решение проблемы отображения более двух измерений на двумерной поверхности. Кора имеет дело по крайней мере с четырьмя наборами значений: двумя для х и у-координат в поле зрения, одним - для ориентации и одним - для различных степеней предпочтения глаза. Две координаты точек на поверхности коры используются для указания позиции поля; две другие переменные удалось разместить в коре благодаря ее подразделению на столь мелкие участки, что, пробежав весь набор ориентации или степеней предпочтения глаза, мы будем иметь лишь такой сдвиг позиций в поле зрения, который будет мал по сравнению с разрешающей силой в этой части зрительного мира. Стратегия членения коры на мелкие вертикальные подразделения явно не ограничена первичной зрительной зоной. Такие подразделения впервые были обнаружены в соматосенсорной области В. Маунткаслом (V. Mountcastle) из Медицинской школы Университета Джонса Гопкинса примерно за 10 лет до нашей работы на зрительной коре. В соматосенсорной области, как мы указывали выше, в основе топографии лежит картирование противоположной половины тела, но сверх этого имеется еще двойная система подразделений, так что есть области, где нейроны реагируют на движение суставов или давление на кожу, и другие области, где нейроны реагируют на прикосновение или отклонение волосков. Как и в случае зрительных колонок, полный комплект (набор нейронов всех типов) занимает здесь место протяженностью около миллиметра. Эти подразделения аналогичны колонкам глазодоминантности в том отношении, что они определяются в первую очередь распределением волокон при входе в кору (там — от левого и правого глаза, здесь — от глубоко расположенных рецепторов и от рецепторов верхних слоев кожи), а не связями внутри коры, типа тех, которые определяют предпочтение ориентации и связанную с этим систему ориентационных колонок. Смысл колонок, обнаруженных в первичной зрительной и соматосенсорной коре, истолкован лучше всего, однако имеются указания на наличие сходных вертикальных подразделений и в некоторых других зонах: ряде высших зрительных областей, сенсорных теменных областях, недавно изученных Маунткаслом, и в слуховой зоне, где Т. Имиг (Т. Imig), X. Эдриен (Н. Adrian) и Дж. Брадж (J. Brugge) из Медицинской школы Висконсинского университета нашли подразделения, в которых два уха представляются попеременно то суммирующими идущую от них информацию, то конкурирующими.
Для большинства из этих физиологически установленных систем (кроме зрительных) до настоящего времени неизвестно анатомических коррелятов. С другой стороны, в последние годы несколько анатомов, в особенности Э. Джонс (Е. Jones) из Медицинской школы Вашингтонского университета, а также Наута (Nauta) и П. Голдмен (P. Goldmen) из Массачусетского технологического института показали, что пути из одной зоны коры в другую (например, из соматосенсорной зоны одной стороны в соответствующую зону на другой стороне) оканчиваются в участках, имеющих правильные чередования с периодом около миллиметра. Здесь колонки видны морфологически, но нет никакой идеи относительно их физиологической интерпретации. Ясно, однако, что тонкие периодические подразделения — действительно общая черта коры мозга. Таким образом, можно сказать, что первое наблюдение Маунткаслом такого свойства способствовало четвертому глубокому прозрению относительно организации коры.
Конечно, было бы неверно считать, что данное рассмотрение зрительной коры в какой-то степени исчерпывает предмет. Кора, по-видимому, имеет дело и с цветом, и с движением, и со стереоскопической глубиной, но в какой степени и каким образом — пока неясно. Из наших работ, относящихся к восприятию глубины, и из работ по цветовому зрению, выполненных С. Зеки (S. Zeki) из Лондонского университетского колледжа, можно заключить, что высшие кортикальные зрительные зоны, в которые первичная кора проецируется непосредственно или обходным путем, могут быть специализированы для обработки соответствующих параметров, но мы еще очень далеки от понимания того, в чем заключается эта обработка.
Что делается за пределами первичной зрительной коры и как информация об ориентации используется на последующих стадиях? Нужно ли думать, что в конечном счете обнаружится клетка, специфически реагирующая лишь на некоторый очень определенный объект? (Обычно в качестве такого объекта выбирают чью-то бабушку по причинам, которые мы уяснить не можем.) Наш ответ состоит в том, что мы сомневаемся в существовании таких клеток, но мы не можем предложить взамен ничего хорошего. К счастью, широкие спекуляции на тему о том, каким образом мозг мог бы работать, это не единственный путь, открытый исследователями. Изучать мозг — это более увлекательное и, кажется, более полезное занятие.
Было время, и не так давно, когда, глядя на миллионы нейронов в различных слоях коры, можно было сомневаться в том, что у кого-нибудь когда-нибудь может возникнуть хоть какая-нибудь идея относительно их функции. Работают ли все они параллельно, как клетки печени или почки, выполняя свои функции сообща, или каждый из них делает что-то свое, особое? Для зрительной коры ответ представляется теперь в общем плане известным: нейроны возбуждаются или тормозятся специфическими стимулами; группы нейронов действительно выполняют специальные преобразования. Если окажется возможным разгадать секреты нескольких подобных областей, будет резонно полагать, что и другие области со временем также раскроют свои тайны.
Э. ЭВАРТС