
Мембраны, через которые необходимо проникнуть веществу для проявления биологической активности, разделяются на несколько типов:
Наиболее простые мембраны, состоящие из липидов и белков – мембраны, перенос через которые осуществляется за счет диффузии.
Диффузия – процесс, при котором частицы (молекулы или ионы) переходят через мембрану из области с высокой концентрацией вещества в область с низкой концент рацией в результате броуновского движения. Процесс может протекать только до выравнивания концетраций, т.е. до достижения равновесия. Для этих мембран характерно, что скорость проникновения вещества через мембрану зависит, как правило, 1) от липофильности соединения и тем выше, чем более жирорастворимо переносимое через мембрану вещество (до определенного предела).
Облегченная диффузия – Для второго типа мембран характерно наличие специфической системы, облегчающей перенос соединения через мембрану, т.е. способствующей диффузии. В этом случае отсутствует также потребление энергии при транспорте. Однако различие с первым типом мембран заключается в том, что возможен облегченный перенос соединений, которые без переносчика проникнуть через мембрану не могут. Например, диффузия молекулы холина через мембрану первого типа практически невозможна из-за наличия тетраалкиламмонийной группы, но специфический переносчик быстро доставляет холин в эритроциты и другие клетки
HO-CH2-CH2-N+(CH3)3 Cl-
Переносчики обладают высокой химической специфичностью, способны узнавать свой субстрат при наличии в среде структурных аналогов.
Т.о. облегченная диффузия – процесс перемещения молекул (по градиенту концентрации) при помощи белков – переносчиков. Специфические переносчики существуют для сахаров, аминокислот, пуриновых и пиримидиновых оснований, нуклеотидов, глицерина и т.д. и т.п. Транспорт этих молекул не может протекать по принципу простой диффузии: проницаемость мембраны для этих соединений стала бы лимитировать скорость обмена веществ. В клетках существует специальный путь переноса таких субстратов с помощью белков-переносчиков, движущей силой для работы которых является разница концентраций ионов натрия, кальция или водорода по обе стороны мембраны. В этом случае транспорт необходимых субстратов в клетку осуществляется совместно с ионами натрия. Таким образом, асимметричное распределение ионов по обе стороны мембраны играет роль дополнительного депо энергии и обеспечивает энергетическую устойчивость метаболизма.
Активный транспорт – это процесс переноса частиц через плазматическую мембрану против градиента концентрации. Он требует притока извне энергии! Активный транспорт ионов против их концентрационного градиента зависит от наличия в клетке АТФ.
АТФ генерируется в организме в результате гликолиза в цитоплазме–бескислородный процесс расщепления глюкозы и поставке компонентов для синтезов органических соединений.
С6Н12О6 → 2 СН3-СО-СООН + 2 АТФ
Гликолизу предшествует подготовительный этап гидролиза крахмала в лизосомах, в пищеварительном канале, сопровождающийся выделением энергии
(С6Н10О5) → С6Н12О6 + Е
Заканчивается кислородным дыханием в митохондриях
СН3-СО-СООН → СО2 + Н2О + 36 АТФ
Суммируя, имеем 1 С6Н12О6 → 74 АТФ
Существует несколько систем активного транспорта ионов в плазматической мембране (ионные насосы): натрий-калиевый насос (Na+/K+ - АТФаза), кальциевый насос (Ca+-АТФаза), водородный насос (Н+-АТФаза).
Одна из наиболее изученных систем активного транспорта – это Na+/K+ - Аденозинтрифосфатаза АТФаза. С ее помощью поддерживаются разные концентрации ионов Na+ и K+ внутри и снаружи клетки. Обычно внутри преобладают иона калия, а снаружи – натрия. Поскольку ионы, хотя и слабо, но все же диффундируют через мембрану, их концентрации стремятся выровняться. Na+/K+ - АТФаза осуществляет активный транспорт, что предотвращает выравнивание концентраций указанных частиц. Na+/K+ - АТФаза – фермент из группы транспортных. Открыт Йенсом Скоу в 1957 г. Он выделил фермент из периферических нервов с помощью уабаина – специфически связывающегося с АТфазой гликозида. В 1997 г. удостоин Нобелевской премии по химии.


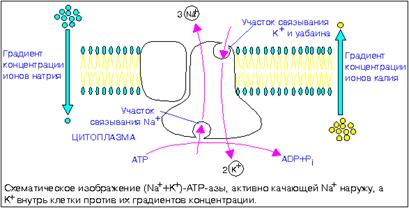
В нейронах мозга, например, на его осуществление затрачивается до 30% всего фонда аденозинтрифосфата, причем спящий мозг расходует на поддержание разницы в распределении одновалентных катионов внутри и вне нейронов почти столько же энергии, сколько и бодрствующий. Использует эту энергию для реализации активного транспорта одновалентных катионов через клеточную мембрану специальный фермент – (Na/K)-активируемая аденозинтрифосфатаза или Na/KATФаза. Она представляет собой сложный белок, встроенный в наружную мембрану клетки и имеющий центры связывания для ионов натрия и калия, а также активный центр, где осуществляются связывание и гидролиз АТФ. Функциональная единица фермента состоит из двух полипептидных цепей: большей (альфа-субъе- диницы) и меньшей (бета-субъединицы), входящих в состав ферментного комплекса в соотношении 1: 1. Меньшая субъединица пересекает мембрану только один раз, в то время как большая – много раз, образуя несколько петель, при этом оба конца пептидной цепи обращены в цитоплазму. Активный центр фермента обращен в цитоплазму и доступен для цитоплазматического АТФ. Центры связывания переносимых ионов локализованы в петле между второй и третьей спиралями, пронизывающими мембрану. Таким образом, альфа-субъединица может выполнять функцию насоса независимо от бета-субъединицы. Однако оба полипептида образуют компактную глобулу, насквозь пронизывающую мембрану. Та часть бета-субъединицы, которая обращена во внеклеточную среду, несет на себе ковалентно присоединенные углеводные фрагменты. По массе и наличию углеводов этот полипептид можно отнести к лектинам – мембранным гликопротеинам, которые отвечают за межклеточное узнавание и адгезию. В процессе белкового синтеза обе субъединицы встраиваются в мембрану одновременно. Существует мнение, что бета-субъединица обеспечивает правильную ориентацию альфа-субъединицы в мембране.
Особая роль Н+-АТФазы заключается в том, что, выкачивая протоны из клетки наружу, она поддерживает рН цитоплазмы близкий к нейтральному (что очень важно для протекания многих ферментативных процессов).
Н+-АТФаза – это интегральный белок, полипептидная цепь которого десять раз пересекает поверхностную (плазматическую) мембрану. Молекулярная масса одной субъединицы фермента ∼104 кД. Полагают, что в мембране Н+-АТФаза функционирует в виде олигомера и состоит из двух субъединиц. Хорошо изучены как первичная структура Н+-АТФазы (то есть аминокислотная последовательность ее полипептидной цепи), так и структуры более высоких порядков (вторичная, В молекуле Н+-АТФазы различают несколько участков (доменов), из которых основные – это домен, связывающий АТФ четырьмя местами связывания, и домен, имеющий отношение к переносу протона, включающий канал. Истинным субстратом Н+-АТФазы является не сама АТФ, а ее комплекс с магнием (Mg-АТФ). В процессе работы Н+-АТФаза подвергается фосфорилированию-дефосфорилированию и обратимо меняет свою конформацию.
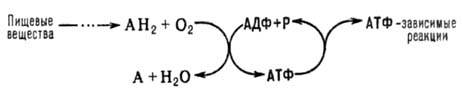
Окислительное фосфорилирование - синтез АТФ из аденозиндифосфата и неорганического фосфата, осуществляющийся в живых клетках, благодаря энергии, выделяющейся при окислении органических веществ в процессе клеточного дыхания. В общем виде окислительное фосфорилирование и его место в обмене веществ можно представить схемой:
При этом она переходит из формы Е1 в форму Е2 и обратно. В форме Е1 она связывает протон на внутренней стороне мембраны, а в форме Е2 освобождает его на наружной стороне. На 1 мкм2 поверхности мембраны приходится 104 молекул Н+-АТФазы. Каждая молекула работает со скоростью от 20 до 100 оборотов в секунду и переносит от 105 до 106 протонов в секунду на 1 мкм2. Как правило, отношение количества перенесенных протонов к количеству гидролизованных молекул АТФ равно единице.
Для нормального функционирования Н+-АТФазы в мембране необходимо присутствие некоторых фосфолипидов. Оптимум рН этой ферментной системы лежит в слабокислой среде (рН 6,3–6,5).
Еще один способ проникновения веществ в клетку – пиноцитоз, который заключается в следующем: небольшие области мембраны образуют «впячивания», которые далее преобразуются в пузырьки и пузырьки оказываются «вне» клетки, если они образовались на внутренней стороне мембраны, и «внутри» клетки, если они возникли на внешней ее поверхности.
Таким образом через мембрану проникают макромолекулы, которые не могут пересечь этот барьер путем диффузии.
Сходен с пиноцитозом фагоцитоз: макромолекулярные комплексы поступают в клетку или выделяются из нее в виде пузырьков, окруженных липидными мембранами. Ферменты либо гормоны как бы выдавливаются из клетки в виде таких пузырьков. Если в клетку попадают чужеродные белки, то в лизосомах они гидролизуются под действием протеолитических ферментов. Бактерии и вирусы попадают в клетку также путем фагоцитоза. В организме человека борьба с таким вторжением осуществляется клетками иммунной системы – макрофагами. Они служат для разрушения проникающих в организм чужеродных белков до того, как последние успевают вызвать повреждение клеток.
Мембраны 4-го типа, имеющие те же свойства, что и мембраны 1-го типа, но отличаются определенным размером (по диаметру). Примером могут служить почечные клубочки, пропускающие любые молекулы, размеры которых (~3 нм) меньше, чем диаметр молекулы альбумина. Альбумин - основной белок крови, вырабатываемый в печени человека. Около 40% альбумина содержится в крови, остальные 60% - в межклеточной жидкости. Проявляет высокую связывающую способность по отношению к различным низкомолекулярным соединениям, содержит гидрофильные и липофильные связи.


Молекула альбумина содержит много дикарбоновых аминокислот, поэтому может удерживать в крови катионы Са2+, Сu2+, Zn2+. Другая особенность альбумина – не содержит углеводных компонентов. Функции альбумина:
- на 80% определяет коллоидно-осмотическое давление плазмы;
- «молекула – такси» переносит билирубин, уробилин, соли желчных кислот, некоторые экзогенные вещества – пенициллин, сульфамиды, ртуть, липид-ные гормоны, некоторые лекарства, такие как варфарин, фенитоин и др.
Для простых мембран, для проникновения через которые не требуется потребление энергии, диффузия крайне затруднительна для ионов, во-первых, потому, что мембрана имеет заряд, вследствие чего разноименные заряды притягиваются, т.е. прилипают к ней, а одноименные отталкиваются от нее, и, во-вторых, потому, что ионы в организме гидратированы, что многократно увеличивает их размеры.
О проникновении лекарственных веществ (вообще ксенобиотиков) в мозг. Диффузия в мозг затруднена больше, чем в другие органы и ткани организма. От посторонних веществ мозг защищен ГЭБ –гемато-энцефалическим барьером, который имеет наиболее плотное перекрывание клеток и, будучи «проходим» для многих липофильных соединений, совершенно непроницаем для ионов. Однако, это в норме. При воспалительных процессах картина меняется и многие вещества уже могут проходить ГЭБ. После преодоления ГЭБ вещество должно еще проникнуть через мембраны внутри мозга и распределиться по его отделам.