Содержания
Введение
- 1 История изучения
- 2 Виды РНК
- 3 Химический состав и модификации мономеров
- 4 Структура
- 5 Сравнение с ДНК
- 6 Синтез
- 7 Типы РНК
- 7.1 Участвующие в трансляции
- 7.2 Участвующие в регуляции генов
- 7.3 В процессинге РНК
- 8 Геномы, состоящие из РНК
- 8.1 РНК-содержащие вирусы
- 8.2 Ретровирусы и ретротранспозоны
- 9 Гипотеза РНК-мира
- 10 Литература
Введение
Рибонуклеи́новые кисло́ты (РНК) — нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания — аденин, цитозин, гуанин и урацил (в отличие от ДНК, содержащей вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусах. Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) подвергаются сплайсингу и принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК. Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах. Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы) у отдельных РНК обнаружена собственная энзиматическая активность, способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами. Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
История изучения
Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus). Позже было обнаружено, что бактериальные клетки, в которых нет ядра, тоже содержат нуклеиновые кислоты. Значение РНК в синтезе белков было предположено в 1939 году в работе Торбьёрна Оскара Касперссона, Жана Брачета и Джека Шульца. Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при её введении в ооциты образуется тот же самый белок. В Советском Союзе в 1956-57 годах проводились работы (А. Белозёрский, А. Спирин, Э. Волкин, Ф. Астрахан) по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляет рибосомальная РНК.Северо Очоа получил Нобелевскую премию по медицине в 1959 году за открытие механизма синтеза РНК. Последовательность 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холея, за что в 1968 году он получил Нобелевскую премию по медицине. В 1967 Карл Вёзе предположил, что РНК обладают каталитическими свойствами. Он выдвинул так называемую Гипотезу РНК-мира, в котором РНК прото-организмов служила и в качестве молекулы хранения информации (сейчас эта роль выполняется в основном ДНК) и молекулы, которая катализировала метаболические реакции (сейчас это делают в основном ферменты). В 1976 Уолтер Фаэрс и его группа в Гентском Университете (Голландия) определили первую последовательность генома РНК-содержащего вируса, бактериофага MS2. В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения. Приблизительно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микроРНК, играют регуляторную роль в онтогенезе нематод C.elegans.
Виды РНК
Молекулы РНК в отличие от ДНК являются однонитевыми структурами. Схема построения РНК аналогична ДНК: основу образует сахарно-фосфатный остов, к которому присоединяются азотистые основания. Различия химического строения заключаются в следующем: дезоксирибоза, присутствующая в ДНК, заменена на молекулу рибозы, а тимин представлен другим пиримидином — урацилом. Молекулы РНК в зависимости от выполняемых функций подразделяются на три основных вида: информационные, или матричные (мРНК), транспортные (тРНК) и рибосомальные (рРНК). В ядре клеток эукариот содержится РНК четвертого вида — гетерогенная ядерная РНК (гяРНК), которая является точной копией соответствующей ДНК.
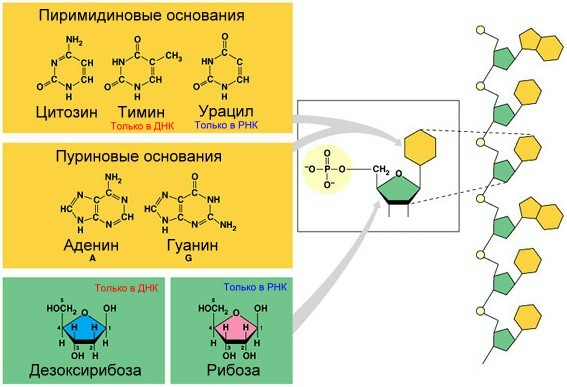
Рис.1. Строение ДНК и РНК
Химический состав и модификации мономеров

Рис.2. Химическое строение полинуклеотида РНК
Нуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1 присоединено одно из оснований: аденин, гуанин, цитозин или урацил. Фосфатная группа соединяет рибозы в цепочку, образуя связи с 3 атомом углерода одной рибозы и в 5 положении другой. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — полианион. РНК транскрибируется как полимер четырёх оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C), но в «зрелой» РНК есть много модифицированных оснований и сахаров. Всего в РНК насчитывается около 100 разных видов модифицированных нуклеозидов, из которых 2-О-метилрибоза наиболее частая модификация сахара, а псевдоуридин — наиболее часто встречающееся модифицированное основание. У псевдоуридина (Ψ) связь между урацилом и рибозой не C — N, а C — C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК. Другое заслуживающее внимания модифицированное основание — гипоксантин, деаминированный гуанин, нуклеозид которого носит название инозина. Инозин играет важную роль в обеспечении вырожденности генетического кода. Роль многих других модификаций не до конца изучена, но в рибосомальной РНК многие пост-транскрипционные модификации находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующим в образовании пептидной связи.
Структура
Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырёх нуклеотидов, в которой есть пара оснований аденин — гуанин.

Рис.3. Разные формы нуклеиновых кислот. На рисунке (слева направо) представлены A (типична для РНК), B (ДНК) и Z (редкая форма ДНК)
Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2 положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК. У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка. Второе последствие наличия 2 гидроксильной группы состоит в том, что конформационно пластичные, то есть не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять.

Рис.4. Вторичная структура РНК-компонента теломеразы простейших
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуемой с помощью водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы. В силу большого числа возможных вариантов спаривания оснований предсказание вторичной структуры РНК — гораздо более сложная задача, чем предсказание вторичной структуры белков, но в настоящее время есть эффективные программы, например, mfold. Примером зависимости функции молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES — структура на 5 конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требующего наличия особого модифицированного основания (кэпа) на 5 конце и белковых факторов инициации. Первоначально IRES были обнаружены в вирусных РНК, но сейчас накапливается всё больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса. Многие типы РНК, например, рРНК и мяРНК в клетке функционируют в виде комплексов с белками, которые ассоцииируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
1. ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
2. Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
3. ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка.
Синтез
Синтез РНК в живой клетке проводится ферментом — РНК-полимеразой. У эукариот разные типы РНК синтезируются разными, специализированными РНК-полимеразами. В целом матрицей синтеза РНК может выступать как ДНК, так и другая молекула РНК. Например, полиовирусы используют РНК-зависимую РНК-полимеразу для репликации своего генетического материала, состоящего из РНК. Но РНК-зависимый синтез РНК, который раньше считался характерным только для вирусов, происходит и в клеточных организмах, в процессе так называемой РНК-интерференции. Как в случае ДНК-зависимой РНК-полимеразы, так и в случае РНК-зависимой РНК-полимеразы фермент присоединяется к промоторной последовательности. Вторичная структура молекулы матрицы расплетается с помощью хеликазной активности полимеразы, которая при движении субстрата в направлении от 5 к 3 концу молекулы синтезирует РНК в 3 --> 5 направлении. Терминатор транскрипции в исходной молекуле определяет окончание синтеза. Многие молекулы РНК синтезируются в качестве молекул-предшественников, которые подвергаются «редактированию» — удалению ненужных частей с помощью РНК-белковых комплексов. Например, у кишечной палочки гены рРНК расположены в составе одного оперона (в rrnB порядок расположения такой: 16S — tRNAGlu 2 — 23S —5S) считываются в виде одной длинной молекулы, которая затем подвергается расщеплению в нескольких участках с образованием сначала пре-рРНК, а затем зрелых молекул рРНК. Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК. После завершения транскрипции РНК часто подвергается модификациям (см. выше), которые зависят от функции, выполняемой данной молекулой. У эукариот процесс «созревания» РНК, то есть её подготовки к синтезу белка, часто включает сплайсинг: удаление некодирующих белок последовательностей (интронов) с помощью рибонуклеопротеида сплайсосомы. Затем к 5 концу молекулы пре-мРНК эукариот добавляется особый модифицированный нуклеотид (кэп), а к 3 концу несколько аденинов, так назваемый «полиА-хвост».

Рис.5. Структура молоточкового (hammerhead) рибозима, который расщепляет РНК
Типы РНК
Матричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка. Однако подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибироваться с отдельных генов (например, рибосомальные РНК) или быть производными интронов. Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, которые участвуют в процессе трансляции. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК. По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
7.1. Участвующие в трансляции
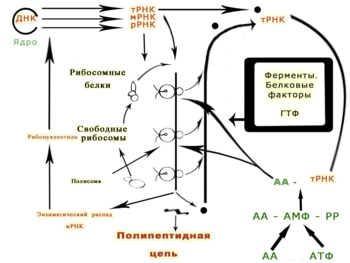
Рис.6. Роль разных типов РНК в синтезе белка (по Уотсону)
Информация о последовательности аминокислот белка содержится в мРНК. Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскирибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК. В безъядерных клетках (бактерии и археи) рибосомы могут присоединяться к мРНК сразу после транскрипции участка РНК. И у эукариот, и у прокариот цикл жизни мРНК завершается её контролируемым разрушением ферментами рибонуклеазами.
Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК.
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки.
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию.
7.2. Участвующие в регуляции генов
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов.
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами. У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов. В качестве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух Х-хромосом у самок млекопитающих. Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5 и 3 нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин.
7.3. В процессинге РНК
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК). Кроме того, интроны могут катализировать собственное вырезание. Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышко и тельцах Кахаля. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК. Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте - особом участке митохондрии протистов-кинетопластид (например, трипаносом).