Расписанные выше превращения (а также все те, что будут описаны далее) реализуются на основе ряда механизмов. Механизмов морфогенеза, общих для всех тканевых зачатков. Привожу их список:
- Пролиферация клеток и их рост
- Детерминация и дифференцировка
- Эмбриональная индукция
- Миграция клеток
- Гибель клеток (апоптоз)
Поговорим о каждом.
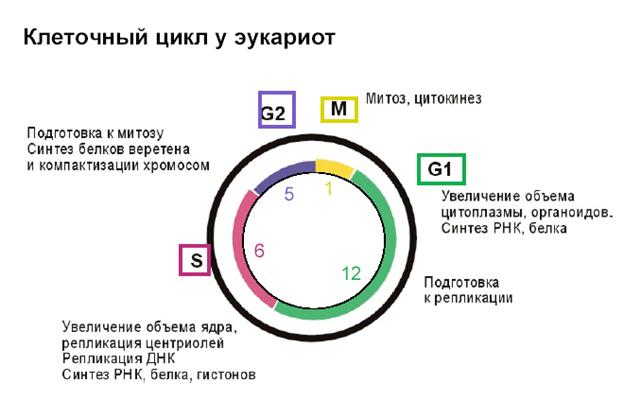
Пролиферация клеток и рост клетки являются разными этапами жизненного цикла клетки. Рост – это прерогатива G0-G1/2 фазы, пролиферация – SиM–фазы. Переход из одной фазы цикла в другую осуществляется циклин – зависимыми протеинкиназами. В определённые фазы клеточного цикла уровень экспрессии каждого из циклинов(регуляторная субъединица ЦЗК) направленно изменяется.
Из G0 – G1 – образование комплексов циклина Д с ЦЗК – 4 или 6
G1 – S – Циклин Е с ЦЗК – 2.
Вход в митоз – ЦЗК – 1 с Циклином В
Выход покоящейся клетки из G0 и вступление в митоз инициируется секретируемыми цитокинами из группы факторов роста, а также в результате взаимодействия клетки (интегринов) с белками внеклеточного матрикс. Эти закономерности действуют как в постнатальной жизни, так и во внутриутробной.
Детерминация клетки. Изначально клетка полипотентная (плюрипотентная), т.е. способна стать практически любой клеткой известного науке фенотипа. Однако, она должна выбрать путь дальнейшего развития, в начала в рамках четырёх основных типов тканей (эпителий, соединительная ткань, мышечная, нервная), а затем – уже внутри этого типа.
Механизмы эмбрионального развития следует объяснить, исходя из представлений о регуляции работы генов, и в первую очередь, это касается детерминации и дифференцировки клеток. Простыми словами, детерминация – это программирование клетки на экспрессию определённого набора генов. А вот реализация этой программы уже составляет суть клеточной дифференцировки.
|
|
Виды детерминации:
• оотипическая – программирует развитие зародыша из зиготы;
• зачатковая – программирует развитие органов и систем из эмбриональных зачатков;
• тканевая – программирует развитие специализированных тканей.
При этом происходит закрепление окончательных свойств, и ткани теряют способность к взаимопревращениям. Такое явление называют стабильной детерминацией. (в опухолевых клетках существует феномен эпителиально – мехенхимального перехода, когда исходный эпителиальный фенотип ткани приобретает черты, характерные для низкодифференцированной соединительной ткани. Это выражение опухолевой прогрессии)
Как происходит дифференцировка клеток?
Изначально задаётся программа реализации определённых генов, и сначала генов, определяющих специфичность типа ткани, затем – отдельной группы внутри типа ткани (внутри нервной, эпителиальной и т.д.). Экспрессия генов задаёт определённый фенотип, характерный для той или иной разновидности ткани. Однако следует упомянуть ещё вот о чём.
Эмбриональная индукция. Определённые тканевые зачатки на разных этапах эмбриогенеза берут на себя функцию индукторов. Эмбриональная индукция заключается в том, что один тканевой зачаток побуждает другую популяцию клеток к детерминации и дифференцировки в том или ином направлении. Это реализуется также с помощью цитокинов, которые воздействуют на соответствующие рецепторы клеток и изменяют транскрипцию генов соответствующим образом.
|
|
Примеры: самый известный пример связан с экспериментом Ганса Шпемана. Тот, со своей сотрудницей Хильдой Мангольд (возможно и любовницей, кто знает), пересадили кусок ткани из дорсальной губы бластопора (отверстия первичной полости тела) в вентральную область эмбриобласта другого тритона. После они смогли различить в тритоне – реципиенте ткани донора, а именно – хорду и миотомы. Таким образом, дорсальная губа бластопора воздействует на определённую группу мезодермальных клеток и заставляет их становиться хордой. Вентральная губа, по разным данным, индуцирует образование нервной трубки из популяции эктодермальных клеток. В целом, эта концепция применима и к более частным моментам, например, образованию различных клеток внутри одной ткани.
Миграция клеток. Наиболее очевидно это прослеживается в отношении мезодермы и мезенхимы. Мезодерма сама является результатом миграции клеток из экто и энтодермы. Мезенхимальные клетки (те самые, что дают начало мышечной ткани, скелетной, хрящевой, эндотелию сосудов, соединительной ткани и проч.), мигрируют из места первичной локализации в место окончательной закладки. За ними следуют нервные волокна, осуществляющие их иннервацию. Следы такой миграции можно проследить уже у родившегося человека: ярким примером является диафрагмальный нерв, который тянется аж с шеи до самой диафрагмы, проходя через всю грудную клетку. Или вагус. Другим примером миграции служит переселение первичных половых клеток. Изначально они расположены в стенке желточного мешка. Одно время даже считали, что именно желточный мешок является местом их образования. Позднее показано, что они оттуда мигрируют в гонады. А сами по себе дифференцируются в гаметоциты ещё на стадии деления бластомеров. По этой причине их нельзя отнести ни к одному зародышевому листку: ни эктодермы, ни энтодермы, ни мезодермы.
Но если же половые клетки не достигли места окончательной локализации, если мезенхимальные клетки не получают иннервации и по целому ряду других причин, клетки подвергаются апоптотической гибели. Связано это с тем, что на определённых этапах те или иные клетки мешают дальнейшему морфогенезу. И от них надо избавляться.