Анатомические характеристики ствола
Ствол мозга (truncus encephali) – является продолжением спинного мозга в полости черепа и в своем строении сохраняет ряд характерных для него черт [1; 629]. Физиологи анатомически объединяет три отдела головного мозга: продолговатый мозг, мост и средний мозг, совместная деятельность которых формирует основные стволовые функции [4; 159].
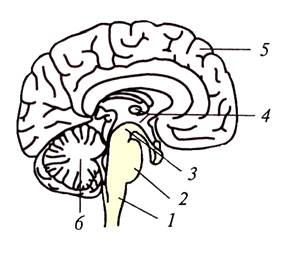
Ствол головного мозга, сагиттальный разрез:
1 – продолговатый мозг;
2 – мост;
3 - средний мозг;
4 – промежуточный мозг (таламус и гипоталамус);
5 – мозжечок.
Продолговатый мозг
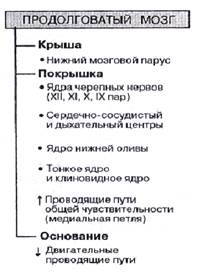
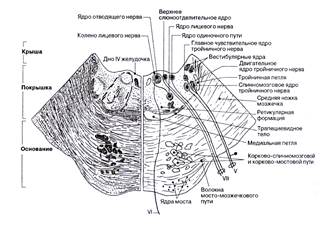
Продолговатый мозг (bulbus, medulla oblongata) – самая каудальная часть ствола мозга, расположен между спинным мозгом и варолиевым мостом. В продолговатом мозгу расположены ядра черепно-мозговых нервов (XII, XI, X, IX пар), разделенные проводящими путями, проходящими через продолговатый мозг, как в восходящем, так и в нисходящем направлении [3; 63].
Так, ядро языкоглоточного нерва (n. glossopharyngeus, IX пара) расположено в продолговатом мозге. Двойное (двигательное) ядро вызывает поднимание глотки и гортани, опускание мягкого нёба и надгортанника в глотательном рефлексе. Чувствительное ядро одиночного пути получает вкусовую, тактильную, температурную, болевую и интероцептивную чувствительность от слизистой оболочки глотки, задней трети языка, барабанной полости и каротидного тельца, входит в состав соответствующих анализаторов, участвует в рефлексах жевания, глотания, в секреторных и моторных пищеварительных рефлексах, а также в сосудистых и сердечных рефлексах (из каротидного тельца). Нижнее слюноотделительное (парасимпатическое) ядро стимулирует секрецию околоушной слюнной железы.
Ядро блуждающего нерва (n. vagus, X пара), состоит из двойного (двигательного) ядра, сокращает мышцы нёба, глотки, верхней части пищевода и гортани, участвует в рефлексах глотания, рвоты, чихания, кашля, в формировании голоса. Чувствительное ядро одиночного пути, иннервируя слизистую оболочку нёба, корня языка, дыхательных путей, аортальное тельце, органы шеи, грудной и брюшной полости, участвует в качестве афферентного звена в глотательном, жевательном, дыхательных, висцеральных рефлексах. Оно входит в проводниковый отдел интероцептивного, вкусового, тактильного, температурного и болевого анализаторов. Заднее (парасимпатическое) ядро, иннервируя сердце, гладкую мускулатуру и железы органов шеи, грудной и брюшной полостей, участвует в сердечных, легочных, бронхиальных, пищеварительных рефлексах.
Двигательное ядро добавочного нерва (n. accessories, XI) посылает импульсы к грудино-ключично-сосцевидной и трапециевидной мышцам, что ведет к их сокращению и вызывает наклон головы набок с поворотом лица в противоположную сторону, поднимание плечевого пояса вверх, сведение лопаток к позвоночнику.
Двигательное ядро подъязычного нерва (n. hypoglossus, XII) иннервируя мышцы языка, вызывает его движение в рефлексах жевания, сосания, глотания и осуществлении речи. Другая информация об иннервации ядер черепных нервов представлена в таблице №1 и на рисунке №1.1 [4; 160 – 161].
Другой характерной структурой продолговатого мозга является ретикулярная формация – скопление нейронов со специфическими свойствами, основная масса которых занимает центральную часть продолговатого мозга. В основном ретикулярная формация на этом уровне ствола имеет нисходящие влияние на моторные спинальные центры. От ретикулярного гигантоклеточного ядра (РГЯ) продолговатого мозга идет частично перекрещенный латеральный ретикулоспинальный путь, волокна которого оканчиваются на вставочных нейронах спинного мозга. Через эти интернейроны они возбуждают ά- и γ – нейроны мышц- сгибателей конечностей и реципрокно тормозят мышцы-разгибатели. На этой схеме показаны основные моторные центры ствола мозга и их пути, регулирующие тонус мышц туловища и конечностей, где КЯ – это красное ядро и руброспинальный путь; РЯМ – ретикулярная ядра моста и медиальный ретикулоспинальный путь; ВЯ – вестибулярные ядра и вестибулоспинальный путь; РГЯ – ретикулярное гигантоклеточное ядро и латеральный ретикулоспинальный путь; МН – спинальные мотонейроны [4; 164 – 165].
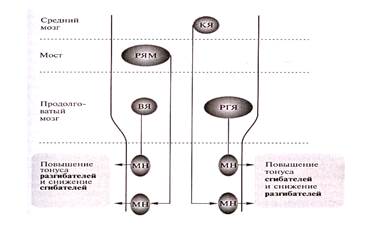
При этом ретикулоспинальные пути, облегчающие активность спинного мозга, берут свое начало от каждого уровня ствола мозга. Пути, тормозящие моторную активность, начинаются преимущественно в бульбарном отделе, и их влияние билатерально. Показано, что ретикулоспинальные пути, начинающиеся от области варолиева моста, тормозят альфа и гамма мотонейроны сгибателей и активируют мотонейроны разгибателей, в то время как волокна от бульбарного отдела оказывают обратное действие. Электрическая стимуляция обширных областей ретикулярной формации ствола мозга приводит к ритмическим движениям, тремору, нередко сопровождающимся тоническими сокращениями, которые оказываются весьма длительными [3; 69 – 70]. В функциональном и топографическом плане латеральный ретикулоспинальный путь сходен с руброспинальным и кортикоспинальным путями; они образуют нисходящую латеральную флексорную (сгибательную) систему. От ретикулярных ядер моста идет неперекрещенный медиальный ретикулоспинальный путь, оканчивающийся на интернейронах спинного мозга. Через них осуществляется стимуляция альфа и гамма нейронов мышц-разгибателей осевой мускулатуры тела (туловища и проксимальных отделов конечностей) и через тормозные интернейроны тормозятся сгибатели. В функциональном и топографическом отношении этот путь сходен с вестибулоспинальными путями, они составляют медиальную нисходящую экстензорную (разгибательную) систему [4; 165 – 166].
«Варолиев мост»
Варолиев мост (pons) – имеет снизу и спереди вид толстого белого валика, расположенного в средней части ствола мозга. Вверху он граничит с ножками мозга, а внизу – с продолговатым мозгом [2; 95]. Проводящие пути в мосту головного мозга идут не общими столбами, а как бы разбиваются на отдельные тонкие пучки, проходящие в узких промежутках между собственными ядрами моста (V, VI, VII пары). Такое расположение является оптимальным для выполнения одной из важнейших функцией моста головного мозга – обеспечивающий обмен информацией между различными проводящими системами, осуществляемого за счет перехода коллатеральных (идущих в обход основного пути) нервных волокон от одного проводящего пучка к другому. Благодаря такой функции моста совершаются мгновенные движения [1; 629].

В каудальной части моста снаружи от латеральной петли локализованы тела нейронов лицевого нерва (n. facialis, VII пара). Двигательное ядро вызывает сокращение мимической и вспомогательной жевательной мускулатуры, регулирует передачу звуковых колебаний в среднем ухе в результате сокращения стременной мышцы. Чувствительное ядро одиночного пути, иннервируя вкусовые луковицы передних ⅔ языка, анализирует вкусовую чувствительность, участвует в моторных и секреторных пищеварительных рефлексах. Верхнее слюноотделительное (парасимпатическое) ядро стимулирует выделение секретов подъязычной, подчелюстных слюнных и слезной желез.
В задней половине моста на дне VI желудочка, у средней линии расположено ядро отводящего нерва (n. abducens, VI пара), которое, сокращая наружную прямую мышцу глаза, вызывает поворот его наружу.
На среднем уровне моста непосредственно впереди ядра лицевого нерва локализованы тела нейронов моторного ядра тройничного нерва (n. trigeminus, V пара), которое имеет двигательное и чувствительные ядра. Двигательное ядро иннервирует жевательную мускулатуру и вызывает движение нижней челюсти вверх, вниз, в стороны и вперед, а также направляет мягкое нёбо и барабанную перепонку. Чувствительные ядра (среднемозговое, мостовое, спинальное) получают от кожи, слизистых оболочек, органов лица и головы тактильную, температурную, висцеральную, проприоцептивную, болевую импульсацию, входят в проводниковый отдел соответствующих анализаторов и участвуют в различных рефлексах, например жевательном, глотательном, чихательном [3; 79 и 4; 160]. Другая информация об иннервации ядер черепных нервов представлена в таблице №1 и на рисунке №1.1
Ретикулярная формация в области моста – является ростральным продолжением ретикулярной формации продолговатого мозга. Основное ретикулярное ядро моста – это каудальное ретикулярное ядро, которое приходит продолжением гигантоклеточного ядра продолговатого мозга. Орально ретикулярное ядро моста, являющееся продолжением каудального ядра и переходящее в ретикулярную формацию среднего мозга, и ретикулярное ядро покрышки моста, расположенное вентральнее каудального ретикулярного ядра моста. Волокна от нейронов ретикулярного ядра покрышки моста проецируются в мозжечок, а от других ретикулярных нейронов моста – в спинальный мозг (ретикулоспинальный путь) к нейронам VIII пластинки по Рекседу; волокна от нейронов каудального ядра моста достигают шейных, грудных и поясничных сегментов спинного мозга. При этом стимуляция ретикулярных ядер моста может вызвать в экстензорных спинальных мотонейронах ответы как с коротким латентным периодом (фазные мотонейроны), так и с длинным (тонические мотонейроны) [3; 79 – 80].
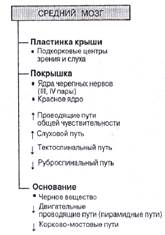
«Средний мозг»
Средний мозг (mesencephalon) – переднюю (нижнюю) часть его составляют ножки мозга, по которым проходят основные нисходящие проводящие пути, а заднюю (верхнюю) – пластинка четверохолмия. В четверохолмии различают два верхних холмика, содержащие ядра первичных зрительных центров, и два нижних, содержащих ядра первичных слуховых центров. Эти центры среднего мозга являются не только пунктом для передачи информации в высшие центры зрения и слуха, расположенные в коре больших полушарий, но и участвуют в организации непроизвольной автоматизированной двигательной реакции на неожиданное звуковое или световое раздражение – так называемых «старт-рефлексов». Полостью среднего мозга является узкий канал – водопровод мозга [1; 629 и 2; 98]. В числе ядер черепно-мозговых нервов среднего мозга можно выделить III, IV пары.
Так ядро глазодвигательного нерва (n. oculomotorius, III пара) расположено в среднем мозге. Двигательное ядро сокращает верхнюю, нижнюю, внутреннюю прямые, нижнюю косую мышцы глаза и мышцу, поднимающую верхнее веко, участвуя в глазодвигательных рефлексах. Добавочное (парасимпатическое) ядро, иннервируя сфинктер зрачка и ресничную мышцу, осуществляет рефлексы сужения зрачка и аккомодации глаза.
Ядро блокового нерва (n. trochlearis, IV пара) иннервируя верхнюю косую мышцу, оно осуществляет поворот глазного яблока вниз и кнаружи [4; 159 – 160].

Кроме глазодвигательного ядра и блокового ядра в среднем мозге имеются – красное ядро и черная субстанция, которые находится непосредственно над ножками мозга. Красное ядро получает по нисходящим путям импульсы от коры мозга, подкорковых двигательных ядер и мозжечка и передают сигналы по руброспинальным путям к нейронам спинного мозга. Кроме того, красное ядро связано с ретикулярной формацией верхней части продолговатого мозга и участвует в регуляции мышечного тонуса; нарушение этой связи приводит к состоянию децеребрационной ригидности. Черная субстанция координирует акты жевания и глотания, участвует также в регуляции пластического тонуса, а у человека – и в мелких движениях пальцев руки [1; 629 и 3; 83].
Ретикулярная формация среднего мозга имеет обширные связи с глубинными отделами полушарий головного мозга и выполняет ряд важнейших функций. Представлена в основном клеточной группой, локализированной кпереди от орального ретикулярного ядра варолиева моста. По нисходящим ретикулоспинальным путям мезенцефальные ретикулоспинальные нейроны могут оказывать на клетки спинного мозга как активирующее, так тормозящие влияние. Возбудительный тонус в ретикулярных нейронах может поддерживаться благодаря циркуляции импульсов по нейронным замкнутым кольцевым цепям, что обусловливает готовность к деятельности этих отделов ЦНС, обеспечивая их быстрое реагирование и включение реакции настораживания.
Одной из важных функций мезенцефалона является его участие в перераспределении мышечного тонуса и осуществлении и координировании тонических рефлексов, в частности, статокинетических и установочных, или выпрямительных, сохраняющихся у мезенцефального животного; в запуске этих рефлексов участвуют рецепторы лабиринтов, шейных мышц и кожной поверхности тела, в осуществлении – красное ядро.
Таким образом, среднемозговой уровень центральной нервной системы обеспечивает и переработку сенсорной информации, и двигательные и диенцефальнокортикального, и бульбарноспинального уровней [1; 629 и 3; 84 – 85].
Стволовые двигательные проводящие пути
Стволовые рефлексы запускаются с рецепторов вестибулярного аппарата, проприорецепторов шеи, рецепторов сетчатки и тактильных рецепторов. Эфферентные влияния из моторных центров ствола распространяются на мускулатуру конечностей и туловища по четырем нисходящим путям:
1) вестибулоспинальному,
2) руброспинальному,
3) латеральному,
4) медиальному ретикулоспинальному.
Вестибулоспинальный путь начинается преимущественно от вестибулярного латерального ядра (а также от верхнего и медиального вестибулярных ядер), получающего афферентную импульсацию от рецепторов отолитового аппарата и ампул полукружных каналов. Его волокно оказывают возбуждающее действие на спинальные альфа и гамма мотонейроны мышц-разгибателей конечностей, туловища, шеи, а также реципрокное тормозящее влияние на мотонейроны сгибателей. Аналогично на мышечный тонус влияет медиальный ретикулоспинальный путь. Вместе они составляют медиальную нисходящую разгибательную систему.
Противоположное действие на мышечный тонус оказывает руброспинальный путь, который начинается от красного ядра покрышки среднего мозга и идет в спинной мозг, возбуждая, альфа и гамма нейроны мышц-сгибателей и реципрокно тормозя мотонейроны мышц-разгибателей. Аналогично на мышечный тонус действуют латеральный ретикулоспинальный и пирамидный кортикоспинальный пути. Вмести они составляют латеральную нисходящую сгибательную систему. Таким образом, в стволе мозга имеются 4 главных парных моторных центра и пути, регулирующие тонус мышц туловища и конечностей [4; 168 – 169].
Рисунок №1.1.
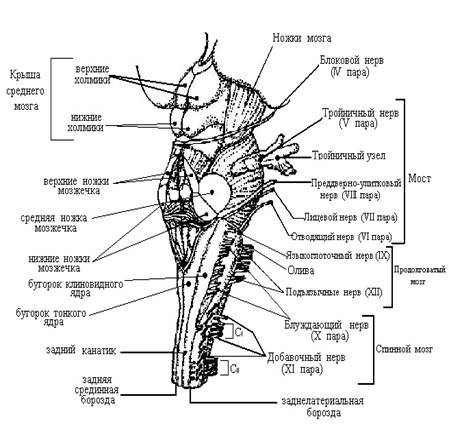
Ствол мозга с ядрами черепных нервов
Список используемой литературы
Популярная медицинская энциклопедия. Гл. ред. В.И. Покровский – 4-е изд., «Книгочей», 1997, 688 с.
Козлов В.И., Цехмистренко Т.А. Анатомия нервной системы / учебное пособие для студентов. – М, 2004. – 208 стр.
Алейникова Т.В. Физиология центральной нервной системы: Учеб. пособие / Т.В. Алейникова, В.Н. Думбай, Г.А. Куратаев, Г.Л. Фельдман – Ростов н/Д: Феникс, 2000. – 384 стр.
Смирнов В.М. Физиология центральной нервной системы: Учеб. пособие для студ. высш. учеб. заведений / В.М. Смирнов, В.Н. Яковлев, В.А. Правдивцев. – 3-е изд., испр. и доп. – М.: «Академия», 2005. – 368 стр.