Кафедра иммунологии
Контрольная работа
По дисциплине: «Радиационная генетика»
Вариант № 5
Выполнила студентка 4 курса
группа А82МЕЭ2
Галустян Эрна Григорьевна
Минск 2021
Особенности индукции облучением точковых и хромосомных мутаций. Связь с репарацией (доказательства).
Термин „точковая мутация” долгое время понимался разными исследователями по-разному, поэтому следует уточнить, что согласно современным представлениям точковыми мутациями мы называем стабильные изменения на уровне нуклеотидов ДНК. При образовании точковой мутации не нарушается целостность хромосомы и не затрагивается ее белковый компонент. Предположение о разной зависимости выхода точковых и хромосомных мутаций от эффективности репарационных систем было высказано нами в 1974г. [Моссэ И.Б., 1974г.] и подтверждено работами последних лет. Данные, свидетельствующие о независимости или слабой зависимости формирования точковых мутаций от интенсивности процессов репарации, получены при исследовании различных факторов.
Радиочувствительность стадий гаметогенеза. В радиобиологии существует установившееся мнение о том, что радиочувствительность разных стадий гаметогенеза различна. Такое явление объясняется разной эффективностью репарационных процессов на разных стадиях. Однако это положение верно лишь для мутаций аберрантного происхождения. Что же касается точковых мутаций, то по этому типу повреждений радиочувствительность разных стадий, как сперматогенеза, так и овогенеза оказывается одинаковой. Так, Лефевр не обнаружил никаких различий в радиочувствительности постмейотических половых клеток дрозофилы по тесту точковых мутаций и пришел к выводу, что радиочувствительность спермиев и сперматид по точковым мутациям одинакова. Наряду с этим выход транслокаций в сперматидах больше, чем в спермиях, в 11,6 раза, а рецессивных сцепленных с полом- летальных мутаций (смесь хромосомных и точковых) – в 3,7 раза.
|
|
Группа японских исследователей при изучении на дрозофиле мутаций dumpy точкового и хромосомного происхождения неоднократно отмечала разницу в изменении частот этих типов мутаций при различных воздействиях. Так, в работах показано, что при облучении выход мутаций dumpy точкового происхождения одинаков на разных стадиях сперматогенеза у дрозофилы, в то время как индукция мутаций dumpy хромосомной природы различается.
Не обнаружено различий между зрелыми и незрелыми ооцитами по чувствительности к индукции облучением мутаций dumpy точковой природы. Установлено, что вариации в общей частоте мутаций на протяжении овогенеза обусловлены мутациями аберрантного происхождения, тогда как выход точковых мутаций мало зависит от фазы овогенеза. Эти данные полностью согласуются с данными
полученными в работе при исследовании индукции мутаций в 10 специфических локусах половой хромосомы в ооцитах и овогониях дрозофилы.
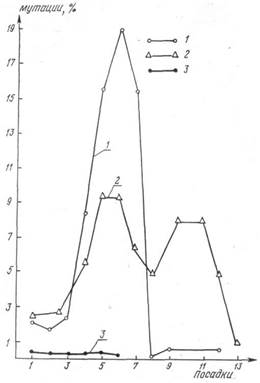
Рис.1 Характер выхода различных типов мутаций, индуцированных облучением: 1 – частота транслокаций;2 – частота рецессивных летальных мутаций; 3 – частота мутаций при цитологически нормальных хромосомах.
Таким образом, по тесту точковых мутаций спермии имеют практически одинаковую радиочувствительность со сперматидами, а овогонии с ооцитами, несмотря на огромную разницу в метаболизме и эффективности репарационных процессов в этих клетках.
|
|
Радиочувствительность разных линий. При изучении разных линий дрозофилы, отличающихся по своей радиочувствительности, показано, что выход точковых мутаций одинаков, а различия обусловлены лишь неодинаковой индукцией хромосомных аберраций [Александров И.Д., 1976г.].
Кислородный эффект. Известно, что кислород, присутствующий во время облучения, подавляет работу репарационных систем. При отсутствии кислородного влияния на выход хромосомных перестроек у линий дрозофилы, дефектных по эксцизионной и репликационной репарации, кислородный эффект является модификацией способности к репарации
Модификация кислородом частоты радиационных мутаций показана лишь для хромосомных аберраций, а индукция облучением точковых мутаций не изменяется при воздействии кислородом или азотом. Например, по данным работы, обработка половых клеток дрозофилы азотом после облучения в кислороде повышает частоту транслокаций в этих клетках, но не изменяет выхода точковых мутаций (рецессивные летальные мутации в кольцевой Х-хромосоме). Аналогично воздействие кислородом после облучения в азоте уменьшает выход аберраций хромосом, но не влияет на индукцию точковых мутаций. При этом если выход одиночных и двойных разрывов при облучении в воздухе, азоте и кислороде различается существенно, то данные по изменению выхода поврежденных оснований при облучении в кислороде отсутствуют.
Выдерживание в кислороде после облучения в азоте способствует репарации повреждений в сперматидах дрозофилы, при этом уменьшается выход индуцированных облучением аберраций, но частота генных мутаций в специфических локусах половой хромосомы и аутосомы II не изменяется.
|
|
Влияние мощности дозы. Выход видимых мутаций и мутаций dumpy у дрозофилы изучался при облучении γ -лучами с разной мощностью дозы – 3000 и 30 рад/мин (Единицы измерений доз облучения в цитируемых работах нами не переведены в единицы СИ, так как не указано, являются ли эти дозы экспозиционными или поглощенными.). Оказалось, что частота мутаций аберрантной природы была значительно ниже при малой мощности дозы, чем при высокой, но выход точковых мутаций остался одинаковым при уровнях мощностей дозы, различающихся в 100 раз. Таким образом, индукция точковых мутаций не зависит от мощности дозы облучения.
Влияние типа излучения. Репарация повреждений, индуцированных нейтронным облучением, не обнаружена. Такие повреждения либо вовсе не репарируются, либо репарируются с большим трудом. При изучении выхода мутации dumpy у дрозофилы установлено, что ОБЭ нейтронов по сравнению с рентгеновским излучением в 2 раза выше для мутаций хромосомной природы, чем для точковых.
Влияние ЭМС. Известно, что ЭМС вызывает преимущественно генные мутации. Выход генных мутаций у дрозофилы под влиянием ЭМС не модифицируется даже кофеином. Показано также, что в клетках китайского хомячка повреждения, индуцированные ЭМС, необратимы и приводят к индукции и экспрессии мутаций в отсутствие синтеза ДНК. Частота генных мутаций в локусе HPRT в клетках китайского хомячка, индуцированных этилнитрозомочевиной, не изменяется в разных послеэкспозиционных условиях, в которых снижается частота СХО. Авторы данной работы пришли к выводу, что повреждения, ведущие к СХО, в отличие от нерепарируемых „мутагенных” изменений репарируются.
Влияние гена, нарушающего рекомбинацию. При исследовании влияния γ-лучей на индукцию мутаций у дефектной по рекомбинации линии дрозофилы c3G было выявлено, что частота точковых мутаций в сперматоцитах мутантной и дикой линий одинакова, а индукция гиперплоидных самцов различна. Автор предполагает, что c3G ген включается в процесс индукции ионизирующей радиацией больших структурных изменений и не связан с индукцией генных мутаций.
Для всех приведенных фактов возможно лишь одно объяснение – отсутствие влияния вообще или слабое влияние репарационных систем на выход точковых мутаций.
Общность происхождения хромосомных
и точковых мутаций
Ряд современных данных свидетельствует о возможной связи между первичными повреждениями азотистых оснований и формированием хромосомных перестроек. Рассмотрим некоторые из них.
1. Неионизирующие излучения и химические мутагены не способны вызывать фосфодиэфирные и межуглеродные разрывы. Для них характерны лишь реакции, обусловливающие потери и модификации оснований. Тем не менее, эти мутагены образуют такие же структурные перестройки хромосом, как и ионизирующие излучения.
2. При введении в пиримидиновое ядро заместителей Br, Cl и при включении в ДНК БДУ возникают спонтанные и увеличивается выход индуцированных УФ и рентгеновским излучением аберраций хромосом. При этом радиочувствительность хроматид зависит от степени замещения тимидина ДНК на БДУ. При замещении более чем на 60 % количество разрывов в облученных хроматидах в 3 раза выше, чем при отсутствии замещения. Установлено, что в бромурацильной ДНК под действием УФ и рентгеновского излучения цепочка реакций начинается с диссоциации атома брома, затем происходят свободнорадикальные превращения сахарного остатка, что приводит в конечном итоге к одиночному разрыву сахарофосфатного остова и образованию структурных мутаций хромосом.
3. Химический распад оснований в ДНК, например, дезаминирование, может содействовать разрыву водородных связей. Кроме того, „стопка” оснований может рассыпаться и без разрыва полинуклеотидной цепи в тех местах, где межплоскостное взаимодействие оснований нарушается вследствие потери или модификации одного из нуклеотидов. Эти факты указывают на то, что разрывы нити ДНК могут быть следствием нестабильности цепи, создавшейся в результате потери или модификации нуклеотидов.
4. К аналогичному выводу приводят и данные о реплицирующейся нестабильности хромосом, при которой разрывы хромосом возникают спустя десятки клеточных поколений после облучения. Следовательно, имеет место репликация предмутационных потенциальных изменений, ведущих к аберрациям хромосом. Отсюда вытекает, что молекулярной сущностью таких потенциальных изменений должны быть изменения нуклеотидной последовательности в ДНК, так как данных о том, что в ДНК может реплицироваться что-либо другое, нет.
5. Ионизирующая радиация может вызвать активацию транспозируемых элементов, находившихся ранее в неактивном состоянии. Транспозоны, меняющие порядок расположения нуклеотидов, повышают темп образования как точковых, так и хромосомных мутаций. Транслокация транспозонов сопровождается крупными делециями и инверсиями.
6. Определенные индуцированные повреждения ДНК -межнитевые сшивки типа димеров тимина – служат причиной образования аберраций хромосом.
Таким образом, можно сделать вывод о том, что при действии облучения на ДНК первичные реакции начинаются с повреждения азотистых оснований. Часть из них (возможно, нерепарабельные или нерепарированные по разным причинам) фиксируются в виде точковых мутаций. Другие могут быть основой для появления структурных мутации хромосом.
Мутационные замены аминокислот бывают двух типов: 1) не нарушающие α- или β-спиральной конформации; 2) приводящие к их повреждению разной тяжести: разрушению N-или С-концов, переходу из α- в β -спиральную конформацию и наоборот или разлому исходной вторичной структуры с образованием двух новых. Может быть, мутационные изменения первого типа, не нарушающие конформации хроматина, не подвергаются воздействию репарационных ферментов и дают начало точковым мутациям, а повреждения второго типа приводят путем различных взаимодействий друг с другом и ферментами репарации к образованию хромосомных перестроек?
В 1980 г. была выдвинута гипотеза, согласно которой в основе инициации хромосомных перестроек лежат нарушения оснований ДНК. Эта гипотеза – итог многолетних исследований механизмов образования аберраций хромосом, в ходе которых было установлено, что при действии репарационных ферментов на поврежденные основания ДНК образуются одиночные разрывы хромосом, которые в свою очередь ферментативно преобразуются в двойные разрывы, дающие начало структурным мутациям.
Примерно 2 % всех повреждений ДНК инициируют хромосомные аберрации. Поскольку в ядре 1% всей ДНК представлено якорной ДНК, предполагается, что молекулярные изменения именно в якорной ДНК являются причиной хромосомного мутагенеза, а повреждения в других участках не приводят к нарушению структурной целостности хромосомы.
Молекулярные аспекты проблемы „горячих точек” в мутагенезе рассматриваются в работе, в которой указывается, что мутации типа замены пар оснований и типа сдвига рамки считывания часто обнаруживаются в участках ДНК с повторами и в асимметричных сайтах квазипалиндромов. Повторы одного или нескольких нуклеотидов являются предпосылкой для смещенного встраивания комплементарных нитей ДНК с образованием свободных от водородных связей петель. Квазипалиндромы имеют крестообразную структуру, в которой сайты асимметрии также создают петли. Мутации, по мнению автора, возникают по местам петель в результате либо эксцизионной репарации, либо аберрантной репликации.
Таким образом, индуцированные облучением точковые мутации формируются в более короткий промежуток времени и в меньшей степени, чем хромосомные перестройки, зависят от различных внутриклеточных процессов, в том числе и репарационных. К тому же в формировании перестроек хромосом существенную роль играет и белок как составная часть хромосомы, что несомненно влияет на репарацию предмутационных изменений.
Сформированные мутации в зависимости от своего функционального значения приводят или к изменению генотипа, или к гибели клетки. Следовательно, если первичные повреждения азотистых оснований не подвергаются воздействию репарационных ферментов (из-за нерепарабельности или недоступности повреждений для этих ферментов), то они фиксируются в виде точковых мутаций. Если же системы восстановления узнают измененное основание и взаимодействуют с ним, то повреждение либо восстанавливается к норме, либо дает начало образованию хромосомной перестройки.
Однако нельзя забывать, что всё это справедливо только для - эукариот, имеющих ДНК, сложно упакованную в составе интерфазных хромосом, и только для ионизирующих излучений, способных повреждать нуклеотидные основания ДНК, не повреждая его надмолекулярной структуры.
Связь с репарацией
Согласно современным представлениям о функционировании репаративных систем в клетках эукариот отдельные биохимические реакции репарации ДНК протекают не независимо, а взаимосвязанно и некоторые из них позволяют клетке функционировать, несмотря на присутствие повреждения, причем синтез ДНК может идти „в обход» нерепарированных повреждений.
Очевидно, этим и объясняются факты отсутствия репарации в клетках млекопитающих, обнаруженные в ряде работ. Так, в работе показано, что в некоторых линиях животных клеток димеры не вырезаются. В работе отмечается, что клетки почти всех млекопитающих вообще лишены фотореактивирующего фермента. В то же время в исследовании делается вывод о том, что в клетках роговицы глаза возникают „не фоторепарируемые поражения”, хотя тесты in vitro (при экстракции ДНК из этих же клеток) показывают, что фотореактивируемые поражения ДНК фактически полностью репарируются.
Нерепарабельные или нерепарированные повреждения доживают до синтеза ДНК и фиксируются в точковые мутации редупликационным механизмом.
Ряд авторов считают, что измененные основания вызывают мутации скорее путем неправильного спаривания, чем путем склонной к ошибкам репарации. Предполагается, что причиной возникновения ошибок спаривания является ионизация оснований ДНК. В работе показано, что частота образующихся ошибочных спариваний оснований не согласуется с таутомерной моделью. Предполагается, что выбор нуклеотидов управляется не ферментом, а различиями в свободных энергиях спаривания оснований, так как отдельные водородные связи обладают различной связывающей способностью в зависимости от их расположения.
Механизм образования замены пар оснований путем неправильного спаривания описан в ряде работ. Например, в работе приводится схема образования и фиксации мутаций путем встраивания измененного основания при вегетативной репликации и делается вывод о том, что фиксация мутаций определенного типа зависит не от эксцизионной репарации, а от вегетативной репликации ДНК. При этом событием, фиксирующим мутации, является ошибка репликации на данном участке ДНК.
Известно несколько типов процессов ошибочного спаривания оснований. Среди них – таутомеризация, ионизация и конформационные изменения (анти -син -вращение пуринов и неоднозначное образование пар оснований). Ряд американских исследователей рассмотрели процесс образования водородных связей между нормальными и модифицированными азотистыми основаниями с новых позиций. Термодинамические исследования ионизации и таутомеризации оснований позволили авторам сделать вывод о важности ионизированных структур для стабилизации модифицированной ДНК в физиологических условиях и о связи между способностью к ошибочному
спариванию измененных мутагеном оснований и образованием ионизированных пар оснований.
На репарацию ДНК у эукариот большое влияние оказывает хроматин. Возможно, из-за этого число генов, контролирующих репарацию у высших организмов, больше, чем у прокариот. По мнению Кимбелла реальные процессы репарации у эукариот еще требуют своего выяснения. Это связано с тем, что ДНК в клетке взаимодействует с различными белками, образуя надмолекулярные структуры высших порядков – хроматин, хромосомы, и эффекты ионизирующей радиации в клетке отличаются от того, что можно ожидать, исходя из опытов по облучению ДНК в.
Таким образом, существуют различия в становлении точковых и структурных мутаций и, в частности, в зависимости выхода этих двух типов мутаций от эффективности репарационных процессов в клетке. Ряд авторов приходят к выводу, что аберрации хромосом и точковые мутации являются следствиями разных типов первичных повреждений и репарационных путей. Различие репарационных путей становления этих двух типов мутаций не вызывает сомнений. Но различны ли первичные повреждения, приводящие к точковым и хромосомным мутациям? Многочисленные экспериментальные данные свидетельствуют об обратном
Список литературы
1. Коничев А.С., Севастьянова Г.А. Молекулярная биология. М., 2005, 397 с.
2. Люин Б. Гены. М.:Изд. Бином. 2012, 896 с.
3. Проблемы и перспективы молекулярной генетики: В 2-х т. Том 2 / Отв. ред. Е.Д. Свердлов. – М.: Наука, Т. 1. 2003 – 2004. Т.2. – 2004. – 330 с.
4. Сингер М., Берг П. Гены и геномы. В 2-х т.: Мир. Т.1. – 373 с. Т.2. – 391 с.1998 г.
5. Мушкамбаров Мушкамбаров Н.Н. Молекулярная Молекулярная биология биология: учеб. пособ. для студ. мед. Вузов / Н.Н. Мушкамбаров, С.Л. Кузнецов. – М.: ООО "Медицинское информационное агенство", 2003. – 544 с.