Метаботропные рецепторы устроены намного сложнее и существует несколько вариантов развития их эффектов. Эффект лиганда будет развиваться спустя некоторое время и в результате сложных процессов внутри клетки. Т.е. взаимодействие лиганда и рецептора приводит к изменениям в форме (конформации) рецептора, что в свою очередь приводит к запуску каскада биохимических реакций, изменяющих активность внутриклеточных белков и, в конечном счете, изменению функционального состояния клетки. Основные типы мембранных рецепторов:
ü рецепторы, связанные с гетеротримерными G-белками (например, рецептор вазопрессина).
ü рецепторы, связанные с тирозинкиназной активностью (например, рецептор инсулина).
Рецепторы, связанные с гетеротримерными G-белками (или система вторичных посредников)
Рецепторы, связанные с G-белками, представляют собой трансмембранные белки. Сайт связывания с лигандом находится вне клетки, домен белка внутриклеточный находится в связи с другим белком - G-белком. А рядом с этим комплексом «рецептор+G-белок» находится другой мембранный белок-фермент, называемый аденилатциклазой.
Схема реализации эффекта при этом следующая:
1. Соединяется рецептор и лиганд на внешней части мембраны.
2. Рецептор меняет свою конформацию (форму), что его активирует.
3. Активация рецептора приводит к тому, что G-белок диссоциирует (разделяется) на субъединицы: α и βγ.
4. βγ-субъединица в свою очередь активирует рядом расположенную аденилатциклазу.
5. Аденилатциклаза начинает превращать аденозинтрифосфат (который находится в гиалоплазме клетки) в циклический аденозинмонофосфат.
6. Циклический аденозинмонофосфат (цАМФ) в свою очередь активирует протеинкиназы, которые фосфорилируют (присоединяют фосфатную группу).
7. Протеинкиназы в свою очередь изменяют активность белков внутри клетки.
8. Информация, с помощью фосфорилированных (активированных) внутриклеточных белков (пострецепторный передатчик), доставляется в генетический аппарат или ключевым ферментам и структурным белкам цитоплазмы. В результате этого и вызывается (возникает) тот конечный эффект (ответ), который «требовал» от клетки лиганд:
¡ изменяется активность соответствующих ферментов или структурных белков цитоплазмы;
¡ изменяется активность соответствующих генов и скорость синтеза ферментов или структурных белков.
Внутри клетки циклический АМФ разрушается фосфодиэстеразой.
Нами на лекции и выше рассмотрен пример с аденилатциклазой, однако в качестве вторичного посредника может выступать гуанилатциклаза. Принцип тот же, отличие в том, что образуется не циклический аденозинмонофосфат, а циклический гуанозинмонофосфат.
Кроме того, G белок в выше описанном примере является Gs белком(s-стимулирующий), однако бывает еще Gi (ингибирующий) белок, блокирующий аденилатциклазу.
Рецепторы, связанные с тирозинкиназной активностью
По структуре это — трансмембранные белки, субъединицы которых связаны дисульфидными мостиками. Сайт связывания с лигандом находится вне клетки. Соединение лиганда приводит, как правило, к объединению нескольких рецепторов вокруг одного лиганда: один лиганд и два рецептора - димеризация рецептора; один лиганд и три рецептора – тримеризация рецептора и т.д.
Дальнейшая схема реализации эффекта при этом следующая:
1. Лиганд соединяется с 2,3 и т.д. рецепторами.
2. Происходит изменение конформации рецептора, его внутриклеточной части.
3. Рецепторы с тирозинкиназной активностью фосфорилируют последующие внутриклеточные белки, часто тоже являющиеся протеинкиназами, и таким образом передают сигнал внутрь клетки.
4. Информация, с помощью фосфорилированных (активированных) внутриклеточных белков (пострецепторный передатчик), доставляется в генетический аппарат или ключевым ферментам и структурным белкам цитоплазмы. В результате этого и вызывается (возникает) тот конечный эффект (ответ), который «требовал» от клетки лиганд:
¡ изменяется активность соответствующих ферментов или структурных белков цитоплазмы;
¡ изменяется активность соответствующих генов и скорость синтеза ферментов или структурных белков.
То есть, соответствующая адаптационная программа на действие сигнальной молекулы включается, и клетка может нормально функционировать в изменившихся условиях.
Облегченная диффузия
Через биологические мембраны кроме воды и ионов путем простой диффузии проникают многие вещества (от этанола до сложных лекарственных препаратов). В то же время даже сравнительно небольшие полярные молекулы, например, гликоли, моносахариды и аминокислоты практически не проникают через мембрану большинства клеток за счет простой диффузии. Их перенос осуществляется путем облегченной диффузии. Облегченной называется диффузия вещества по градиенту его концентрации, которая осуществляется при участии особых белковых молекул-переносчиков.
Транспорт Na+, K+, Сl-, Li+, Ca2+, НСО3- и Н+ могут также осуществлять специфические переносчики. Характерными чертами этого вида мембранного транспорта являются высокая по сравнению с простой диффузией скорость переноса вещества, зависимость от строения его молекул, насыщаемость, конкуренция и чувствительность к специфическим ингибиторам — соединениям, угнетающим облегченную диффузию.
Все перечисленные черты облегченной диффузии являются результатом специфичности белков-переносчиков и ограниченным их количеством в мембране. При достижении определенной концентрации переносимого вещества, когда все переносчики заняты транспортируемыми молекулами или ионами, дальнейшее ее увеличение не приведет к возрастанию числа переносимых частиц — явление насыщения. Вещества, сходные по строению молекул и транспортируемые одним и тем же переносчиком, будут конкурировать за переносчик — явление конкуренции.
Различают несколько видов транспорта веществ посредством облегченной диффузии
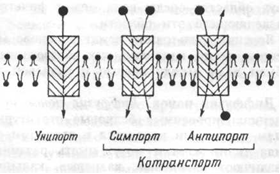
Классификация способов переноса через мембрану.
Унипорт, когда молекулы или ионы переносятся через мебрану независимо от наличия или переноса других соединений (транспорт глюкозы, аминокислот через базальную мембрану эпителиоцитов);
Симпорт, при котором их перенос осуществляется одновременно и однонаправленно с другими соединениями (натрий- зависимый транспорт Сахаров и аминокислот Na+ K+, 2Cl- и котран-спорт);
Антипорт — (транспорт вещества обусловлен одновременным и противоложно направленным транспортом другого соединения или иона (Na+/Ca2+, Na+/H+ Сl-/НСО3- — обмены).
Симпорт и антипорт — это виды котранспорта, при которых скорость переноса контролируется всеми участниками транспортного процесса.
Природа белков-переносчиков неизвестна. По принципу действия они делятся на два типа. Переносчики первого типа совершают челночные движения через мембрану, а второго — встраиваются в мембрану, образуя канал. Промоделировать их действие можно с помощью антибиотиков-ионофоров, переносчиком щелочных металлов. Так, один из них — (валиномицин) — действует как истинный переносчик, переправляющий калий через мембрану. Молекулы же грамицидина А, другого ионофора, встаиваются в мембрану друг за другом, формируя «канал» для ионов натрия.
Большинство клеток обладают системой облегченной диффузии. Однако перечень метаболитов, переносимых с помощью такого механизма, довольно ограничен. В основном, это сахара, аминокислоты и некоторые ионы. Соединения, являющиеся промежуточными продуктами обмена (фосфорилированные сахара, продукты метаболизма аминокислот, макроэрги), не транспортируются с помощью этой системы. Таким образом, облегченная диффузия служит для переноса тех молекул, которые клетка получает из окружающей среды. Исключением является транспорт органических молекул через эпителий, который будет рассмотрен отдельно.