ЯДРО
Ядро – важнейший компонент ядерных (эукариотических) клеток, представляет собой информационный центр клетки, обеспечивающий направленный и регулируемый поток информации. На основе данного потока осуществляются основные внутриклеточные метаболические процессы и деление клетки. В прокариотических клетках ядра нет, а генетический аппарат называется нуклеоидом (нуклеоид – подобный ядру – от «нуклеус» - ядро, «-оид» - подобный). Нуклеоид прокариот – это часть цитоплазмы, в которой находится кольцо двухцепочной (2 цепи) спирали ДНК, без белков-гистонов.
Функции ядра:
1) хранение генетической информации в виде хроматина;
2) воспроизведение генетической информации в процессе репликации ДНК;
3) обеспечение передачи генетической информации потомству;
4) регуляция всех процессов обмена веществ в клетке;
5) реализация генетической информации базируется на матричных процессах: репликации (синтезе ДНК в ядре), транскрипции (синтезе РНК в ядре) и трансляции (синтезе белка в цитоплазме);
6) восстановление генетической информации основанной на репарации ДНК;
7) в ядре образуются субъединицы рибосом;
Строение ядра:
I. Поверхностный аппарат ядра (ПАЯ) – кариотека, кариолемма:
1) наружная ядерная мембрана (НЯМ);
2) внутренняя ядерная мембрана (ВЯМ);
3) перинуклеарное пространство;
4) ядерные поры с поровым комплексом;
5) периферическая плотная пластинка (ППП) – ламина.
II. Ядерный матрикс (ЯМ) -белковый ядерный остов, ядерный скелет:
1) периферический ЯМ: ППП - ламина;
2) внутренний ЯМ: фибриллы кариоскелета (интерхроматиновые и ядрышковые).
III. Кариоплазма (нуклеоплазма):
1) кариолимфа - ядерный сок;
2) фибриллы кариоскелета.
IV. Хроматин – ДНП (Дезоксирибопротеин) = ДНК + протеин (гистоны H1,
H2, H2B, H3, H4 + негистоновые кислые ядерные белки).
1) эухроматин: деспирализован, декомпактизован и функционально активен;
2) гетерохроматин:
а) конститутативный – структурный гетерохроматин (постоянно компактизован);
б) факультативный – функциональный гетерохроматин (м.б. компактизован и не компактизован).
Пример: Х-хромосома (тельце Барра*).
V. Ядрышко:
1. Ядрышковый организатор – это участок ДНК, содержит гены, которые кодируют РНК; в компактной хромосоме соответствует вторичной перетяжке; в ядрышковый организатор клеток человека включены 13, 14, 15, 21, 22 хромосомы.
2. Фибриллярный компонент - РНП фибр. = РНК + белок.
3. Глобулярный компонент - незрелые субъединицы рибосом.
Тельце Барра – компактизованная структура Х-хромосомы (тельце полового хроматина). Причины возникновения тельца Барра: согласно сохранению дозового баланса генов в генотипе для формирования нормального фенотипа у гомогаметного пола происходит инактивация одной из половой хромосомы (ХХ→ХО). Это приводит дозу активно функционирующих Х-генов у данного пола в соответствие с их дозой у гетерогаметного пола ХО или ХУ.
I. Поверхностный аппарат ядра
Поверхностный аппарат ядра (кариолемма) представлен двумя мембранами (наружной и внутренней). Между ними находится перинуклеарное пространство. Обе мембраны имеют жидкостно-мозаичное строение и различаются по набору мембранных белков. Среди этих белков имеются ферменты, переносчики и рецепторы. Главная функция ПАЯ – изоляция гиалоплазмы от кариоплазмы. При этом специальные белки ядерных мембран осуществляют транспортную функцию. В некоторых районах кариолеммы наружная и внутренняя мембраны сливаются и образуют поры. Через эти поры осуществляется связь гиалоплазмы и кариоплазмы. Для регуляции такой связи в порах находятся поровые комплексы.
|





Рис. 1. Строение ПАК.
1. Наружная ядерная мембрана (НЯМ) является продолжением мембран ЭПС. В результате, содержимое полостей ЭПС свободно попадает в перинуклеарное пространство. На внешней стороне НЯМ могут находиться рибосомы.
2. Внутренняя ядерная мембрана (ВЯМ) отличается по составу от НЯМ, прежде всего отсутствием холестерина, обладает меньшей проницаемостью.
3. Перинуклеарное пространство содержит жидкость сходную по составу с гиалоплазмой.
4. Ядерные поры – участки слияния наружной и внутренней мембран кариолеммы в которых находятся поровые комплексы; занимают 3-35% поверхности ядерной оболочки. ПАЯ в клетках животных и человека содержит 2000-4000 поровых комплексов.

|
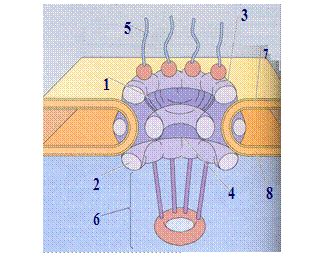
Рис. 2. Структура порового комплекса.
Типичный поровый комплекс (ПК) представляет собой сложную белковую структуру – содержит более 1000 молекул белка. По обе стороны поры (к наружной и внутренней мембранам) в области поры располагаются по 8 белковых глобул (диаметр около 25 нм). Они взаимодействуют между собой и образуют кольцевые структуры диаметром около 120 нм. Одно поровое кольцо располагается с наружной стороны поры (в области основной гиалоплазмы), другое – с внутренней (в области кариоплазмы). Эти периферические глобулы (всего 16) фиксируются в мембранах специальным интегральным белком. От этих глобул к центру сходятся белковые фибриллы, где прикрепляются к центральной глобуле, тем самым формируя перегородку – диафрагму поры, толщиной около 5 нм. С периферическими глобулами связаны тонкие белковые фибриллы, локализованные как в основной гиалоплазме, так и в кариоплазме. Особенностью центральной глобулы является наличие в ней канала диаметром до 15 нм.
Главная функция ПК – транспорт определенных биополимеров из ядра в цитоплазму и из цитоплазмы в ядро. Ионы мелкие и средние органические молекулы, а также
| |||
| |||

|
Наружные глобулы обладают рецепторами к белкам, подлежащим к поступлению из цитоплазмы в ядро. Такие полипептиды называют нуклеофидьными белками. Нуклеофильные белки имеют специальный сигнальный пептид, т.е. белки без него не транспортируются через ПК в ядро. Благодаря этому ПК осуществляет не только транспорт, но и сегрегацию белков, необходимых для функционирования именно в ядерном аппарате.
В ходе транспорта нуклеофильные белки взаимодействуют с тонкими фибриллами гиалоплазмы, связанными с периферическими глобулами наружного кольца ПК, и перемещаются к нему. В центральной глобуле ПК имеются причальные белки (белки-рецепторы), связывающие сигнальный пептид белка. Это взаимодействие индуцирует АТФ-зависимый перенос нуклеофильного полипептида через канал центральной глобулы в кариоплазму. Таким образом, в данном случае центральная глобула выполняет функцию транслокационного комплекса аналогичного таковому в шЭПС.
В ядро через ядерные поры поступают: 1) негистоновые белки (белки-ферменты, которые участвуют в процессах репликации и репарации (восстановление повреждений в ДНК); белки-ферменты, участвующие в транскрипции; белки-репрессоры, которые регулируют процесс транскрипции; белки, входящие в состав субъединиц рибосом; белки ядерного матрикса, образующие кариоскелет); 2) белки-гистоны, которые связываются с молекулой ДНК и образуют хроматин; 3) нуклеотиды; 4) ионы минеральных солей, в частности, ионы Са2+ и Mg2+.
Глобулы внутреннего кольца содержат рецепторы к т-РНК, в периферических глобулах имеется особый белок – переносчик молекул т-РНК. Считается, что сначала транспортируемая молекула взаимодействует с фибриллами периферическиъх гранул в кариоплазме. С их помощью макромолекулы передвигаются к периферическим гранулам, взаимодействуют с рецепторами и транспортируются переносчиками в гиалоплазму. С помощью периферических глобул осуществляется транспорт также других макромолекул из ядра в гиалоплазму. В этих процессах участвуют специальные белки периферических глобул – нуклеопорины.
В центре порового комплекса локализуется центральная глобула. Она связана с периферическими глобулами тонкими фибриллами. Центральная глобула специализируется на транспорте и-РНК из ядра в гиалоплазму. В составе этой глобулы имеются ферменты, которые участвуют в химической модификации и-РНК – ее процессинге, в частности, наблюдается взаимодействие РНК со специфическими белками. Кроме того, через ПК из ядра выводятся и рРНК, но уже в составе субъединиц рибосом, т.е. в комплексе с рибосомальными белками. Считается, что при этом субъединицы рибосом, проходя через ПК, вытесняют центральную глобулу, которая затем возвращается в ПК.
Гранулы поровых комплексов структурно связаны с белками ядерной ламины, которая участвует в их организации. Нуклеоплазма при обмене веществ непосредственно не контактирует с гиалоплазмой.
Обмен веществ между ядром и цитоплазмой осуществляется двумя основными способами:
1) через многочисленные поры, которыми пронизана ядерная оболочка, происходит регулируемый транспорт веществ в ядро и из ядра. Все белки, поступающие из цитоплазмы в ядро, имеют специальную ядерную последовательность, состоящую из нескольких аминокислот, которая опознается белками-рецепторами поровых комплесов;
2) вещества из ядра в цитоплазму и из цитоплазмы в ядро попадают путем отшнуровывания выростов и выпячиваний ядерной оболочки.
Функции комплекса ядерной поры:
1. Обеспечение регуляции избирательного транспорта веществ между цитоплазмой и ядром.
2. Активный перенос в ядро белков, имеющих особую маркировку в виде так называемой последовательности ядерной локализации N uclear L ocalization S equence (NLS), распознаваемой рецепторами NLS (в комплексе поры). Через канал центральной глобулы в ядро попадают определенные белки. Они имеют специальный сигнальный участок, который распознается рецептором центральной глобулы. Белки без такого сигнального участка в ядро не попадают.
3. Перенос в цитоплазму субъединиц рибосом, которые, однако, слишком велики для свободного прохождения через пору; их транспорт, вероятно, сопровождается изменением конформации порового комплекса.
Импорт белков через поровый комплекс ядра включает 5 последовательных этапов:
Белок-импортин ГДФ – гуанозиндифосфат

 ТранспортируемыйNLS-по-
ТранспортируемыйNLS-по-


|
 белокследовательность
белокследовательность
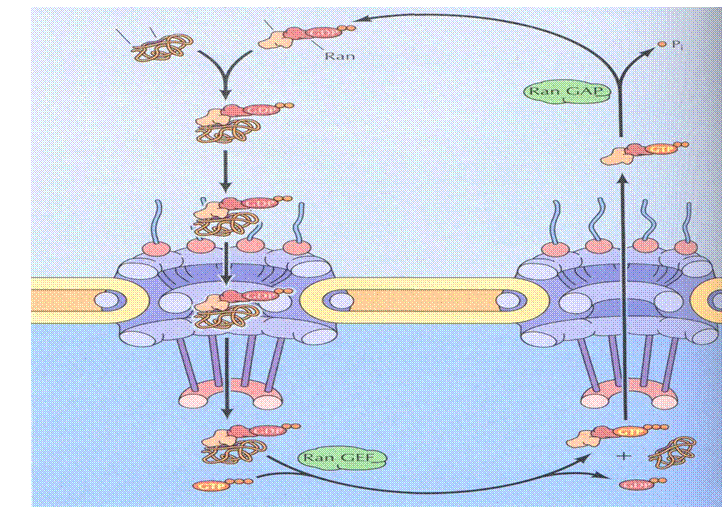 Рис. 4. Импорт белков через ядерную пору.
Рис. 4. Импорт белков через ядерную пору.
1. Распознавание транспортируемого белка, имеющего сигнал ядерной локализации, комплексом белка импортина с белком Ran, связывающим ГДФ*.
2.Связывание образующегося белкового комплекса с белками цитоплазматических филаментов порового комплекса.
3. Перенос белкового комплекса, включающего транспортируемый белок - импортин и белок Ran-ГДФ, через центральный канал порового комплекса.
4. Ферментативное замещение, связанного с белком Ran ГДФнаГТФ* и освобождение транспортируемого белка из комплекса.
5. Перенос комплекса импортин-Ran-ГТФ через ядерный поровый комплекс с последующим ферментативным гидролизом ГТФ до ГДФ (дефосфорилирование).
В основе переноса кариофильных белков через ядерную мембрану лежит различие концентраций гуанозин дифосфата (ГДФ) и гуанозин трифосфата (ГТФ), связанного с белком Ran, по обе стороны ядерной мембраны. Это обусловлено тем, что на наружной ядерной мембране, обращенной к цитозолю, локализуются ферменты, осуществляющие гидролиз гуанозин трифосфата, связанного с белком Rаn, до гуанозин дифосфата, а на внутренней мембране – ферменты, замещающие, связанный с белком Rаn ГДФ до ГТФ.
5. Периферическая плотная пластинка (ППП), или ламина (lamina) – слой толщиной 80 – 300 нм, прилегает изнутри к внутренней ядерной мембране, исключая области пор. Внутренняя ядерная мембрана гладкая, ее интегральные белки связаны с ламиной (периферической плотной пластинкой). Таким образом, ламина является элементом поверхностного аппарата ядра, объединяющим его с ядерным матриксом. Ламина состоит из специальных переплетенных белков – ламинов, образующих периферический кариоскелет в виде ортогональной сетчатой структуры. Белки-ламины относятся к классу промежуточных филаментов (скелетных фибрилл), другие типы которых представлены в цитоплазме и поверхностном аппарате клетки. У млекопитающих известно 3 вида этих белков – это ламины А, В, С. Эти белки поступают в ядро из цитоплазмы. Ламины разных видов взаимодействуют между собой и образуют белковую сеть под внутренней мембраной ядерной оболочки. С помощью ламина В периферическая плотная пластинка соединяется со специальным интегральным белком ядерной оболочки. Этот белок является рецептором ламина В и обеспечивает структурную связь с ППП с ядерной оболочкой. С ППП взаимодействуют и белки периферических глобул «внутреннего кольца» порового комплекса. Ламины А и С образуют сеть тонких фибрилл. К ламину А прикрепляются теломерные участки хромосом.
Функции ламины:
1. Ламина поддерживает форму ядра (формообразующая функция). Даже если обе ядерные мембраны разрушить, то ядро за счет ламины сохраняет свою форму и поровые комплексы остаются на своих местах.
2. Ламина служит компонентом кариоскелета (скелета ядра). Более того, в районе пор ламины взаимодействуют с цитоплазматическими скелетными фибриллами. В результате формируется структурная связь ламины с опорными элементами поверхностного аппарата клетки. Это обеспечивает единство всех скелетных структур клетки.
3. Ламина участвует в сборке ядерной оболочки (формировании кариолеммы).
4. В интерфазном ядре к ламине А прикрепляется хроматин. Таким образом, ламина обеспечивает функцию фиксации хроматина в ядре (обеспечивает упорядоченную укладку хроматина, участвует в пространственной организации хроматина в интерфазном ядре). Это, в свою очередь, увеличивает скорость и эффективность протекания матричных процессов (репликации и транскрипции) и подготовки клетки к делению.
5. Ламина обеспечивает структурную организацию поровых комплексов.
Итак, изобразить схематично строения ядерного аппарата (а)
и поровых комплексов (б, в) можно следующим образом (рис. 5):
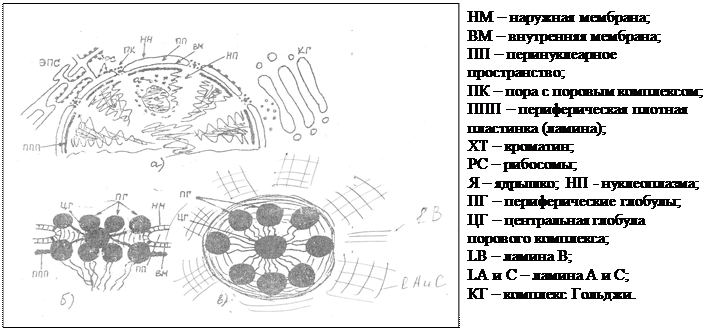
Хотя ламина и представляет собой жесткую скелетную структуру, она может дезинтегрироваться. В частности, это происходит при переходе клетки к процессу деления и связано с активацией специальных протеинкиназ.

Рис. 6. Схема разрушения ламины во время митоза.
Фосфорилирование ламинов с помощью протеинкиназ приводит к разборке ППП. При этом ламин В остается в мембране кариолеммы и мембранных пузырьков, образующихся из ядерной оболочки на первых стадиях деления. Дефосфорилирование ламинов, катализируемое протеинфосфатазами, индуцирует взаимодействие ламинов и формирование ППП в поверхностном аппарате ядра новых клеток.
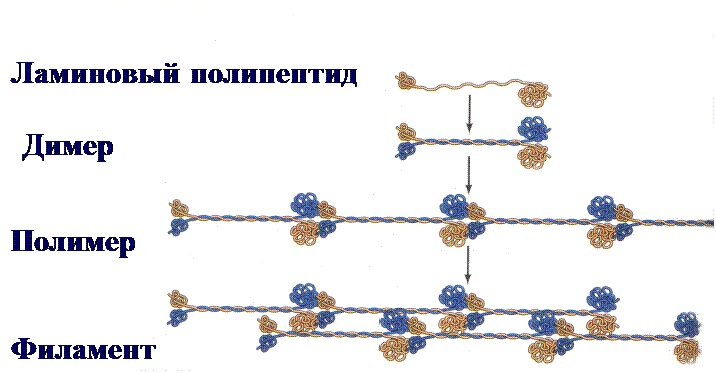
Рис. 7. Схема формирования ламины во время телофазы митоза.
Ламиновые полипепетиды образуют димеры с центральным альфа-спиральным участком, состоящим из двух полипептидных цепей, закрученных друг вокруг друга. Димеры соединяются «голова к хвосту» и образуют полимер. Полимеры, соединяясь друг с другом «бок в бок», формируют филаменты.
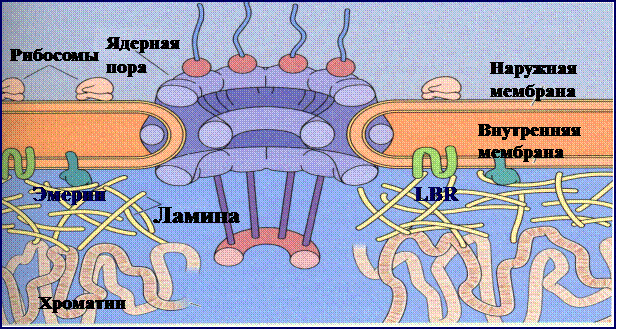
Рис. 8. Связь ламины с различными структурами клетки
II. Ядерный матрикс
Ядерный матрикс (ЯМ) представляет собой систему особых фибриллярных белков толщиной 2-3 нм, локализованных в кариоплазме. ЯМ является универсальным белковым компонентом ядерного аппарата эукариотических клеток. Его универсальность определяется тем, что он обеспечивает оптимальную пространственную организацию и функционирование генетического материала. В составе ЯМ выделяют периферический ЯМ и внутренний ЯМ.
1. Периферический ЯМ представляет собой ламину, т.е. один из элементов ПАЯ, что указывает на структурное единство данных компонентов ядра (рис. 8).
2. Внутренний ЯМ структурно связан с периферическим (ламиной) и представляет собой сложную систему белковых фибрилл, расположенную во всем объеме кариоплазмы. Большинство белков ЯМ относится к группе кислых белков. Внутренний ЯМ дифференцирован на 2 зоны: интерхроматиновую сеть ЯМ и ядрышковую сеть ЯМ.
2.1. Интерхроматиновая сеть ЯМ по составу белков очень разнообразен. В частности в здесь обнаруживаются актиновые микрофибриллы. Одна из функций ЯМ заключается в том, что он представляет собой элемент кариоскелета. Это определяется структурным взаимодействием белковых фибрилл внутреннего ЯМ с ламинами периферического ЯМ (ламиной) и образованием целостной скелетной системы ядра, определяющей его форму. Кроме того, интерхроматиновая сеть фиксируется на внутренних участках хромосом, которые в свою очередь теломерами (концевыми участками) прикрепляются к ламине.
2.2. Одна из частей ядерного матрикса является ядрышковым матриксом. Ядрышковый матрикс состоит из плотно упакованных фибрилл и гранул. Белки ядрышковой сети (фибриллы) взаимодействуют с участками хромосом (вторичными перетяжками) содержащими множество копий генов рибосомальных РНК (саттелитная ДНК). Такие участки получили название ядрышкового организатора, который вместе с ядрышковой сетью и созревающими субъединицами рибосом (гранулы) формируют специфическую структуру ядра – ядрышко.
Таким образом, фибриллы ядерного матрикса контактируют с ламинами ППП и образуют единую систему кариоскелета. Кариоскелет поддерживает форму ядра и обеспечивает определенное расположение хромосом в пространстве, благодаря чему длинные тонкие нити хроматина не спутываются между собой.
III. Кариоплазма
Кариоплазма (нуклеоплазма или ядерный сок), представляет собой желеобразный раствор, в котором находятся как низкомолекулярные (например, нуклеотиды), так и высокомолекулярные (РНК и разнообразные белки, включая ферменты) вещества, а также ионы. Химический состав ее в основном сходен с составом гиалоплазмы, хотя имеются некоторые отличия. В кариоплазму погружены хроматин и ядрышко. Кроме того, в ней располагается ядерный матрикс. Кариоплазма создает специфическую для ядерных структур микросреду и тем самым обеспечивает нормальное функционирование ядерного аппарата. Кариоплазма связана с гиалоплазмой через систему поровых комплексов и транспортных комплексов ядерной оболочки.