Схема строения проводящей системы сердца.
1 - синусовый узел; 2 – атриовентрикулярный узел; 3 – пучок Гиса; 4 –правая ножка пучка Гиса; 5 – общий ствол левой ножки пучка Гиса; 6 – передняя ветвь левой ножки; 7 –задняя ветвь левой ножки; 8 – конечные разветвления ножек пучка Гиса и волокна Пуркинье.
На уровне атриовентрикулярного узла волна возбуждения значительно задерживается. Это обусловлено электрофизиологическими особенностями проводящей ткани атриовентрикулярного узла. Скорость проведения возбуждения по ней варьирует от 5 до 20 см/с. Между атриовентрикулярным узлом и пучком Гиса нет четкой границы.
Прохождение возбуждения по атриовентрикулярному узлу длится в среднем 0,08 с. Вследствие замедленной проводимости и продолжительного рефрактерного периода атриовентрикулярный узел выполняет свою основную функцию – фильтрует подходящие к нему импульсы. Кроме того, он задерживает проведение импульсов. Это создает возможность для окончания возбуждения и сокращения предсердий до того, как начнется возбуждение желудочков.
Атриовентрикулярное соединение обладает функцией автоматизма, вырабатывая 40 – 60 импульсов в 1 мин. Следует отметить, что автоматизм атриовентрикулярного узла не доказан, однако установлено наличие центров автоматизма в нижних отделах предсердия и в пучке Гиса. Клетки водителя ритма в предсердиях, атриовентрикулярном узле и в пучке Гиса называют автоматическими центрами второго порядка. Скорость проведения импульсов в пучке Гиса составляет 1 м/с. Пучок Гиса разделяется сначала на 2 ножки – правую и левую ножки пучка Гиса, причем левая ножка короче правой. Затем пучок Гиса образует 3 ветви: правую ножку и 2 ветви левой ножки пучка Гиса. Эти ветви спускаются вниз по обеим сторонам межжелудочковой перегородки.
Правая ножка, являющаяся продолжением пучка Гиса, представляет собой изолированный тонкий пучок, который проходит по правой стороне межжелудочковой перегородки и направляется к мышце правого желудочка.
Что касается левой ножки, то считают, что сначала она проходит единым коротким стволом по левой половине межжелудочковой перегородки, а затем делится на переднюю (передневерхнюю) и заднюю (задненижнюю) ветви, причем сначала от ветвящейся части пучка Гиса отходит задняя ветвь, а затем – передняя.
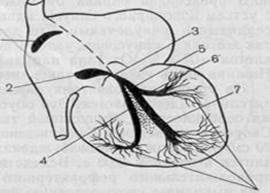
Фронтальный разрез сердца.
Строение внутрижелудочковой проводящей системы. 1 – правая ножка; 2 – правый желудочек; 3 – задняя ветвь левой ножки; 4 – межжелудочковая перегородка; 5 – левый желудочек; 6 – передняя ветвь левой ножки; 7 – левая ножка; 8 – пучок Гиса.

Скорость распространения возбуждения в ветвях и ножках пучка Гиса составляет 3 – 4 м/с. Ножки пучка Гиса и их разветвления, а также конечная часть пучка Гиса обладают функцией автоматизма. Это автоматические центры третьего порядка. Их автоматизм составляет 15 – 40 импульсов в 1 мин.
Ножки пучка Гиса и их ветви состоят из двух видов клеток – Пуркинье и клеток, напоминающих по форме клетки сократительного миокарда.
Поступающий по ним импульс вызывает возбуждение и сокращение миокарда желудочков сердца. Скорость распространения возбуждения по волокнам Пуркинье и миокарду желудочков составляет 4 – 5 м/с. Волокна Пуркинье, по-видимому, также обладают функцией автоматизма (автоматический центр третьего порядка): их автоматизм – 15 – 30 импульсов в 1 мин. В миокарде желудочков волна возбуждения вначале охватывает межжелудочковую перегородку, а затем распространяется на оба желудочка сердца.
В желудочках процесс возбуждения идет от эндокарда к их эпикарду. При возбуждении миокарда создается электродвижущая сила (ЭДС), которая распространяется на поверхность человеческого тела и служит основой для регистрации ЭКГ.
Таким образом, в сердце имеется множество клеток, обладающих функцией автоматизма. Они расположены в синусовом узле, атриовентрикулярном соединении, пучке Гиса и его ножках, а также в желудочках. Однако в норме существует только один водитель ритма, дающий импульсы для возбуждения всего сердца.
W. Einthoven, один из основоположников электрокардиографии, рассматривал сердце как точечный источник электрического тока, расположенный в центре треугольника, образованного правой и левой рукой и левой ногой. При этом образуются точечные положительный и отрицательный заряды, равные по величине, которые представляют собой диполь. Он сделал допущение, что человеческое тело является проводником электрического тока с одинаковым сопротивлением во всех участках. Левая рука, правая рука и левая нога принимались им за три равноудаленные друг от друга и от центра треугольника точки. W. Einthoven считал, что эти точки лежат на вершинах равностороннего треугольника и расположены во фронтальной плоскости. Возникающий при возбуждении сердца вектор ЭДС, по его мнению, смещался также только во фронтальной плоскости. В дальнейшем эта теория была подвергнута существенной коррекции.
Электрическое поле сердца постоянно меняет величину и направление. Это связано с распространением волны возбуждения по мышце сердца. Кроме того, проводящая среда – человеческое тело – неоднородна, что обусловлено разным сопротивлением различных тканей.
Многочисленные исследования показали применимость теории диполя в клинической электрокардиографии. Эта теория позволяет изучать распределение ЭДС сердца не только во фронтальной, но и в других плоскостях.
Для измерения величины потенциала в различных точках электрического поля используют гальванометры. Примером может служить обычный электрокардиограф. ЭДС измеряют с помощью 2 электродов, которые присоединяют к положительному и отрицательному полюсам гальванометра.
Различают активные, или дифферентные, электроды и неактивные, или индифферентные. Активные электроды присоединяют к положительному полюсу гальванометра, индифферентный – к отрицательной клемме гальванометра. Индифферентный электрод теоретически имеет заряд, величина которого приближается к нулю. Примером может служить электрод, расположенный на большом расстоянии от электрического поля, например условно помещенный в бесконечность. Активный электрод измеряет преимущественно потенциалы той точки электрического поля, в которую он помещен. Если активный электрод ставят в точку, расположенную на положительной половине электрического поля, то он измеряет величину положительного потенциала в этой области. Наоборот, если этот электрод помещают в одну из точек отрицательного поля, он измеряет величину отрицательного потенциала в этой области.
Гальванометры устроены таким образом, что если к активному электроду обращен положительный заряд, то гальванометр регистрирует подъем кривой от изолинии, или положительный зубец. Если к активному электроду обращен отрицательный заряд, то гальванометр записывает снижение кривой от изолинии, или отрицательный зубец.
Наличие на поверхности тела человека различных потенциалов позволяет также зарегистрировать разность потенциалов между двумя точками. Для этого надо взять два электрода, поместить их в заданные точки и соединить с гальванометром. В этом случае используются так называемые двухполюсные отведения. Один электрод соединяют при этом с положительным полюсом гальванометра, а другой – с отрицательным.
Гальванометр всегда регистрирует разность потенциалов между двумя электродами. Такая разность возникает не только тогда, когда один участок заряжен положительно, а другой отрицательно, но и в том случае, если оба участка заряжены положительно или отрицательно, но один из зарядов превалирует над другим. Точно так же гальванометр регистрирует разность потенциалов, если один из участков заряжен положительно или отрицательно, а потенциал другого участка равен нулю.
Механизмы аритмий
К основным механизмам аритмий относятся:
1.Аномальный автоматизм.
2.Постдеполяризационная и триггерная активность.
3.Повторный вход и циркуляция волны возбуждения (re-entry).
Автоматизмом называют способность специализированных клеток миокарда спонтанно вырабатывать электрические импульсы. При этом в здоровом сердце существует четкая иерархия с подчинением СА узлу всех нижележащих водителей ритма. При нарушении метаболизма в клетках проводящей системы могут появляться очаги аномального автоматизма, в которых частота генерации импульсов превышает таковую клеток СА узла. Как правило, автоматический центр обладает собственной активностью, которая возникает независимо от активности СА узла, но может модифицироваться последней. Появление аномального автоматизма связано с частичной деполяризацией мембраны в состоянии покоя (фаза 4 ПД) до уровня –40-60 мВ и возникновением входящего кальциевого тока, что способствует дальнейшей деполяризации мембраны до порогового уровня и генерации одиночного ПД или серии потенциалов. Это провоцирует возникновение аритмий, например, очаговых предсердных и желудочковых тахикардий или парасистолии.
Для автоматических тахикардий характерно наличие периодов "разогрева" и "охлаждения" ритма, т.е. постепенное увеличение и уменьшение ЧСС в начале и конце эпизода аритмии. Эти нарушения ритма обычно не купируются электрической стимуляцией во время электрофизиологического исследования (ЭФИ). Однако, в некоторых случаях преждевременные экстрастимулы, попадающие в определенную фазу автоматического цикла, могут вызвать аннигиляцию аномального водителя ритма, т.е. его устранение с внезапным прекращением выработки импульсов. Наличие аномальных центров генерации электрических импульсов можно зафиксировать при проведении внутрисердечного ЭФИ.
Постдеполяризационная и триггерная активность возникает в результате появления во время ПД, индуцированного СА узлом, спонтанных колебаний величины трансмембранного потенциала - ранних или поздних постдеполяризаций. Постдеполяризации могут появиться как реакция на гиперкатехоламинемию, гипокалиемию, гипокальциемию, ацидоз, ишемию, перерастяжение миокарда и другие патологические воздействия. Ранние постдеполяризации возникают обычно на фоне уменьшения частоты основного ритма во время 3 фазы ПД, когда в результате реполяризации мембраны трансмебранный потенциал достигает –50-70 мВ. В этот момент медленные кальциевые каналы могут перейти в состояние активации с повторной деполяризацией мембраны до уровня генерации нового ПД.
Поздние постдеполяризации появляются в 4 фазе ПД в результате гиперполяризации клеточной мембраны, которая может быть индуцирована адреналином или сердечными гликозидами. Постдеполяризации зависят от внутриклеточной концентрации ионов Са2+. В тех случаях, когда амплитуда таких подпороговых осцилляций возрастает и достигает порогового значения генерируется новый ПД или серия ПД. Этот механизм лежит в основе аритмий, возникающих при интоксикации дигиталисом (экстрасистолия, двунаправленная веретенообразная желудочковая тахикардия - ЖТ), реперфузионной ЖТ и аритмий у больных с удлиненным интервалом QT.
В основе подавляющего большинства аритмий лежит механизм риентри (re-entry) – кругового движения электрического импульса. Различают макро- и микрориентри. При макрориентри циркуляция электрического импульса происходит вокруг анатомически невозбудимого препятствия, микрориентри имеет меньшие размеры и может изменять свое местонахождение в миокарде.
Для циркуляции импульса необходимы следующие условия:
1.Два приблизительно параллельных проводящих пути должны соединяться проксимально и дистально, формируя электрический контур.
2.Один из этих путей должен иметь более длинный рефрактерный период, чем другой, что обеспечивает возникновение однонаправленной блокады проведения в одном из сегментов петли re-entry.
3.Длина движущейся волны возбуждения должна быть короче длины петли, благодаря чему перед фронтом волны возбуждения находится участок миокарда, вышедшего из состояния рефрактерности.
При макрориентри импульс, достигший электрического контура, из-за невозбудимости в одном из путей (длительный рефрактерный период) медленно распространяется только по второму пути. К тому времени, когда он достигает соединения между путями, первый путь уже выходит из состояния рефрактерности, и по нему импульс движется в обратном направлении (ретроградно). Далее он вновь достигает медленного пути проведения с коротким рефрактерным периодом и движется по нему антероградно. Циркуляция импульса повторяется, вызывая пароксизм реципрокной (круговой) тахикардии. В результате регулярного выхода импульса из кольца риентри с частотой, значительно превышающей частоту синусового ритма, круг риентри становится патологическим водителем ритма.
При микрориентри отсутствие анатомически невозбудимого субстрата компенсируется наличием зоны абсолютной рефрактерности в центре кольца, которая появляется как результат воздействия импульсов, поступающих с разных сторон при циркуляции возбуждения по окружности. Такой круг с циркулирующим импульсом, получивший название ведущего круга, может перемещаться по миокарду. Радиус его мал и может меняться в зависимости от продолжительности функционального рефрактерного периода, поэтому перед фронтом волны возбуждения отсутствует сколько-нибудь длинный участок, полностью восстановивший свою возбудимость и "голова" волны следует непосредственно за ее "хвостом". В цепи макрориентри, длина которой зависит от размера анатомического препятствия, напротив, имеется "окно возбудимости", длина которого может составлять до 20% от длины всей петли (рис.5).
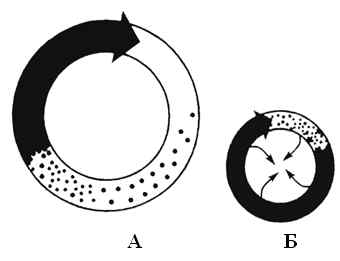
Рис.3 А - макрориентри вокруг анатомического препятствия; Б – микрориентри по механизму ведущего круга. Черным цветом обозначена зона абсолютной рефрактерности, точками – зона относительной рефрактерности, за которой находится возбудимый миокард ("окно возбудимости"). По Кушаковскому М.С. Аритмии сердца.
С механизмом риентри связано возникновение фибрилляции и трепетания предсердий и желудочков (фибрилляция развивается по типу микрориентри, а трепетание – макрориентри), экстрасистолии, реципрокных комплексов и многих разновидностей пароксизмальных тахикардий, в том числе у больных с синдромом WPW и продольной диссоциацией АВ соединения.