Мозжечок
Головной мозг защищён костным футляром мозгового отдела черепа. Мозг имеет овоидную форму благодаря выступающим лобному и затылочному полюсам. Структура головного мозга представлена несколькими отделами: стволом, продолговатым мозгом, мозжечком, варолиевым мостом, средним мозгом и корой больших полушарий. Продольная щель, проходящая по срединной линии мозга, разделяет его правое и левое полушария – гемисферы. Под затылочным полюсом большого мозга пролегает поперечная щель, отделяющая мозжечок – центр координации движений.
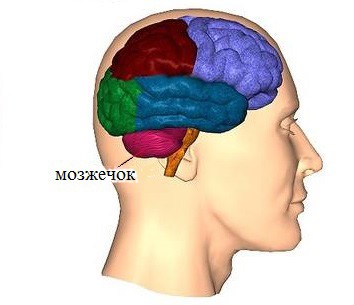
Строение и функции мозжечка
Место расположения мозжечка – задняя черепная ямка. Кпереди от него находятся мост и продолговатый мозг. Мозжечок разделён на 2 полушария, в каждом из которых есть верхняя и нижняя поверхности. Средняя часть мозжечка – червь, разделяет полушария между собой. Кору мозжечка составляет серое вещество тел нервных клеток (нейронов). Кора делится на дольки посредством глубоких борозд, а борозды помельче разделяют между собой листки мозжечка. Кора разветвляется и проникает в тело мозжечка, состоящее из белого вещества. Отростки нейронов представлены в извилинах белым веществом пластинок. Самые нижние дольки, расположенные над большим затылочным отверстием черепа, называются миндалинами мозжечка.
В глубине мозжечка есть парные ядра, состоящие из серого вещества. Эта структура – ядро шатра, относится к вестибулярному аппарату. По бокам от шатра расположены шаровидное и пробковидное ядра, координирующие работу мышц туловища, а также зубчатое ядро, управляющее работой конечностей. С периферией мозжечок связан через другие отделы мозга 3-мя парами ножек. Верхние ножки мозжечка идут к среднему мозгу, средние ножки – к мосту, а нижние – к продолговатому мозгу.
Функции мозжечка в организме человека – координация движений, участие в регуляции работы внутренних органов и скелетных мускулов.
Эмбриональное развитие
Центр координации развивается из нейроэктодермы заднего мозгового пузыря. В конце 8-й недели беременности крыловидные пластинки мозговой трубки эмбриона в области заднего мозга соединяются между собой. На 3-м месяце уже сформированный червь мозжечка имеет 3–4 извилины, разделённые бороздами. К середине 4-го месяца выделяются извилины полушарий мозжечка. На 5-м месяце мозжечок плода уже вполне сформирован. За оставшееся время внутриутробного развития увеличивается его размер, число и глубина борозд и бороздок, разделяющих основные доли на меньшие дольки. К моменту родов мозжечок ребёнка приобретает характерную складчатость и сложность строения.
Симптомы поражения мозжечка
При повреждениях мозжечка нарушаются согласованная работа скелетных мускулов, координация произвольных движений и удержание тела в равновесии.
Мозжечковые расстройства движений имеют характерные признаки:
потеря плавности движений рук и ног;
дрожание в конце целенаправленного движения – интенционный тремор;
изменение почерка;
скандированная речь, которая отличается ритмической, а не смысловой расстановкой ударений в словах;
замедление произвольных движений и речи.
Мозжечковые нарушения равновесия выражаются в головокружениях и расстройстве походки – атаксии. Мозжечковая атаксия похожа на походку пьяного, характерно пошатывание больного в сторону очага поражения. Нарушения движений глазодвигательных мускулов проявляются нистагмом – ритмическим подёргиванием глазных яблок при отведении взгляда в крайние положения. Рассогласование работы мышц конечностей и туловища проявляется также при попытках больного подняться из лежачего положения и сесть без помощи рук.
Мозжечковая атаксия наблюдается при многих заболеваниях и поражениях нервной системы человека: опухолях задней черепной ямки, воспалении мозга и его оболочек, отравлениях, наследственных генетических дефектах, кровоизлияниях различного происхождения.
Врождённые заболевания
Наследственная мозжечковая атаксия Мари – врождённое генетическое заболевание доминантного типа. Болезнь проявляется постепенно нарастающим нарушением координации движений. Отмечается гипоплазия (недоразвитие) мозжечка и его связей с периферией. Характерно начало заболевания в возрасте от 20 до 45 лет с нарушения походки. Постепенно нарастает дрожание в руках, мышечные подёргивания, речь становится скандированной и замедленной. Затем добавляются другие симптомы: птоз (опущение век), падение остроты зрения, нистагм, атрофия зрительных нервов. Болезнь часто сопровождается постепенным снижением интеллекта, нарушением памяти. Инфекционные воспаления, отравления, физические и психические перегрузки способствуют обострениям процесса.
Существуют ещё несколько вариантов хронической атрофии мозжечковой системы: семейная атаксия Фридрейха, торсионная дистопия и другие болезни. При наследственных формах мозжечковой атаксии применяют консервативное лечение, которое уменьшает тяжесть симптомов, улучшает кровоснабжение и питание нервных клеток.
Приобретённые заболевания
Опухоли мозжечка могут быть представлены следующими типами – астроцитома, ангиоретикулома, медуллобластома, саркома. Термин «рак» неприменим к новообразованиям отделов мозга, поскольку в нервной ткани отсутствуют железы – источник роста раковых клеток. Среди злокачественных опухолей самыми частыми являются медуллобластомы и саркомы. Возможно поражение мозжечка метастазами опухолей других органов – меланомы, злокачественных болезней крови.
Черепно-мозговая травма может привести к повреждениям мозжечка, сдавлению его кровоизлиянием – травматической гематомой. При установлении диагноза кровоизлияния проводится хирургическая операция – удаление гематомы.
Причиной кровоизлияния может быть также инсульт – инфаркт мозжечка, возникший вследствие атеросклероза кровеносных сосудов или гипертонического криза. В результате рассасывания мелких кровоизлияний в мозжечке образуются кисты – дефекты нервной ткани, наполненные жидкостью. Функции погибших нервных клеток частично восполняют оставшиеся нейроны.
Точный диагноз очаговых поражений любых отделов мозга устанавливается с помощью магнитно-резонансной томографии (МРТ). Оперативное лечение заболеваний мозжечка проводится при опухолях, очаговых нагноениях (абсцессах), кровоизлияниях, травматических повреждениях.
О трансплантации
Трансплантация мозга и его отделов в настоящее время невозможна по этическим соображениям. Смерть человека констатируется по факту смерти его мозга. Пока есть признаки функционирования мозга, его хозяин считается живым и не может быть донором органов.
22 Базальные ядра. Роль в двигательной активности
Базальные, или подкорковые, ядра представляют собой структуры переднего мозга, к которым относятся: хвостатое ядро, скорлупа, бледный шар и субталамическое ядро. Они располагаются под корой больших полушарий.
Развитие и клеточное строение хвостатого ядра и скорлупы одинаковы, поэтому их рассматривают как единое образование — полосатое тело. Базальные ядра имеют множественные афферентные и эфферентные связи с корой, промежуточным и средним мозгом, лимбической системой и мозжечком. В связи с этим они принимают участие в регуляции двигательной активности и, в частности, медленных или червеобразных движений. Примером таких двигательных актов является медленная ходьба, перешагивание через препятствия и т.д.
Опыты с разрушением полосатого тела доказали его важную роль в организации поведения животных.
Бледный шар является центром сложных двигательных реакций и участвует в обеспечении правильного распределения мышечного тонуса.
Свои функции бледный шар осуществляет опосредованно через образования среднего мозга — красное ядро и черную субстанцию.
Бледный шар также имеет связь с ретикулярной формацией. Он обеспечивает сложные двигательные реакции организма и некоторые вегетативные реакции. Стимуляция бледного шара вызывает активацию центра голода и пищевого поведения. Разрушение бледного шара способствует развитию сонливости и затруднению выработки новых условных рефлексов.
23 Двигательные зоны коры головного мозга
Двигательная область коры головного мозга: расположение, строение и функции
Часть мозга, которая выполняет работу произвольных движений является двигательной (моторной) корой. Необходимые движения осуществляются таким образом, чтобы они лучше всего подходят для текущей позиции индивида.
Левая двигательная кора участвует в регулировании движения правой стороны тела, а правая двигательная область коры головного мозга координирует движения левой стороны тела.
Задумывались ли вы, какая часть мозга играет решающую роль в обеспечении произвольных движений? Это двигательная кора. Она расположена в задней части лобной доли ― 1 из 4 основных долей, передней части мозга. Двигательная кора лежит перед центральной бороздой, как структура, которая позволяет отличить лобную долю и теменной доли.
Функция
Как следует из названия, двигательная кора отвечает за двигательные функции тела, она принимает участие в движении мышц для выполнения определенных задач. Наша способность производить движения, такие как собирание мелких предметов, привязаны к нормальной работе двигательной коры.
Будь то небольшие движения, такие как перемещение пальцев или большие движения, такие, как ходьба и бег, двигательная кора играет решающую роль прямо от начала выполнения движения. Проще говоря, сокращение мышц для нормального движения - это то, что находится под контролем двигательной коры.
В момент начала движения, верхние двигательные нейроны, расположенные в двигательной коре, передают электрические сигналы, которые идут через другие структуры мозга, такие как базальные ганглии и мозговой ствол, и поступают на нижние моторные нейроны, расположенные в спинном мозге. Сигнал затем проходит вниз от нижнего моторного нейрона к другому и, наконец, прибывает на нужные группы мышц. Получив сигнал, мышцы сокращаются, вызывая произвольные движения.
Премоторная кора
Для того, чтобы выполнить определенное движение, определенные мышцы должны сужаться. Премоторная кора головного мозга анализирует и точно определяет группы мышц, которые должны быть использованы для выполнения определенных движений. Она учитывает текущее положение и позу тела при выборе набора мышц для выполнения движения. Например, пиная футбольный мяч, премоторная кора решает какие мышцы ног задействовать, чтобы инициировать действие.
Первичная моторная кора
Эта часть мозга непосредственно взаимодействует с двигательными нейронами спинного мозга, чтобы стимулировать предназначенные мышцы. Короче говоря, первичная моторная кора участвует в передаче сигналов, которые вносят непосредственный вклад в создание движения.
Дополнительная моторная область
Одна из основных функций деятельности ДМО является надзор за ручной координацией. В ручной координации, множественные движения должны быть выполнены одновременно для достижения желаемого действия или задачи.
Например, действия, которые предполагают использование обеих рук потребуют координации между левой и правой двигательной корой. Это делается ДМО, которая позволяет производить совмещение движений одновременно для выполнения сложных действий. Поэтому наша способность двигать одновременно обе руки для выполнения определенной задачи приписывается работу ДМО.
ДМО также помогает определять последовательность движений, так как для выработки оптимального пути движения, необходимо осуществлять комплекс действий.
Сразу отмечу еще один момент заключается в том, что двигательная кора не решает, выполнять ли движение или нет. Например, увидев кусочек шоколада, лежащего на полу, решение его забрать принимается префронтальной корой, которая лежит непосредственно перед премоторной. Префронтальная кора, соответственно, дает сигналы двигательной области коры головного мозга, чтобы осуществлять движение. Кроме того, двигательная кора не единственная область, которая участвует в выполнении точных движений. Другие части мозга, такие как базальные ганглии и мозжечок, в равной степени способствуют производству движений.
24 Кора больших полушарий, ее участие в инициации и поддержании активности
По особенностям клеточного состава поверхность коры подразделяют на структурные единицы следующего порядка: зоны, области, подобласти, поля.
Зоны коры головного мозга разделяются на первичные, вторичные и третичные проекционные зоны. В них расположены специализированные нервные клетки, к которым поступают импульсы от определенных рецепторов (слуховых, зрительных и т.д.). Вторичные зоны представляют собой периферические отделы ядер анализаторов. Третичные зоны получают обработанную информацию от первичных и вторичных зон коры больших полушарий и играют важную роль в регуляции условных рефлексов.
В сером веществе коры больших полушарий различают сенсорные, моторные и ассоциативные зоны:
o сенсорные зоны коры больших полушарий - участки коры, в которых располагаются центральные отделы анализаторов:
зрительная зона — затылочная доля коры больших полушарий;
слуховая зона — височная доля коры больших полушарий;
зона вкусовых ощущений — теменная доля коры больших полушарий;
зона обонятельных ощущений — гиппокамп и височная доля коры больших полушарий.
Соматосенсорная зона находится в задней центральной извилине, сюда приходят нервные импульсы от проприорецепторов мышц, сухожилий, суставов и импульсы от температурных, тактильных и других рецепторов кожи;
o моторные зоны коры больших полушарии - участки коры, при раздражении которых появляются двигательные реакции. Располагаются в передней центральной извилине. При ее поражении наблюдаются значительные нарушения движения. Пути, по которым импульсы идут от больших полушарий к мышцам, образуют перекрест, поэтому при раздражении моторной зоны правой стороны коры возникает сокращение мышц левой стороны тела;
o ассоциативные зоны - отделы коры, находящиеся рядом с сенсорными зонами. Нервные импульсы, поступающие в сенсорные зоны, приводят к возбуждению ассоциативных зон. Особенностью их является то, что возбуждение может возникать при поступлении импульсов от различных рецепторов. Разрушение ассоциативных зон приводит к серьезным нарушениям обучения и памяти.
Речевая функция связана с сенсорными и двигательными зонами. Двигательный центр речи (центр Брока) находится в нижней части левой лобной доли, при его разрушении нарушается речевая артикуляция; при этом больной понимает речь, но сам говорить не может.
Слуховой центр речи (центр Вернике) расположен в левой височной доле коры больших полушарий, при его разрушении наступает словесная глухота: больной может говорить, излагать устно свои мысли, но не понимает чужой речи; слух сохранен, но больной не узнает слов, нарушается письменная речь.
Речевые функции, связанные с письменной речью — чтение, письмо, — регулируются зрительным центром речи, расположенным на границе теменной, височной и затылочной долей коры головного мозга. Его поражение приводит к невозможности чтения и письма.
В височной доле находится центр, отвечающий за запоминание слое. Больной с поражением этого участка не помнит названия предметов, ему необходимо подсказывать нужные слова. Забыв название предмета, больной помнит его назначение, свойства, поэтому долго описывает их качества, рассказывает, что делают с этим предметом, но назвать его не может. Например, вместо слова «галстук» больной говорит: «это то, что надевают на шею и завязывают специальным узлом, чтобы было красиво, когда идут в гости».
Функции лобной доли:
o управление врожденными поведенческими реакциями при помощи накопленного опыта;
o согласование внешних и внутренних мотиваций поведения;
o разработка стратегии поведения и программы действия;
o мыслительные особенности личности.
В кору больших полушарий, к сенсорным нейронам поступают импульсы от всех рецепторов организма через ядра таламуса. И каждый орган имеет свою проекцию или корковое представительство, расположенное в определенных областях больших полушарий.
В коре больших полушарий имеется четыре чувствительные и четыре двигательные области.
Нейроны двигательной коры получают афферентную импульсацию через таламус от мышечных, суставных и кожных рецепторов. Основные эфферентные связи двигательной коры осуществляются через пирамидные и экстрапирамидные пути.
У животных наиболее развита лобная область коры и ее нейроны участвуют в обеспечении целенаправленного поведения. Если удалить эту долю коры, животное становится вялым, сонливым. В височной области локализуется участок слуховой рецепции, и сюда поступают нервные импульсы от рецепторов улитки внутреннего уха. Область зрительной рецепции находится в затылочных долях коры головного мозга.
Теменная область, внеядерная зона, играет важную роль в организации сложных форм высшей нервной деятельности. Здесь расположены рассеянные элементы зрительного и кожного анализаторов, осуществляется межанализаторный синтез.
Рядом с проекционными зонами располагаются ассоциативные зоны, которые осуществляют связь между сенсорной и двигательной зонами. Ассоциативная кора принимает участие в конвергенции различных сенсорных возбуждений, позволяющей осуществлять сложную обработку информации о внешней и внутренней среде.
25 Теория уровней управления движениями
Бернштейн Н.А. обнаружил, что в зависимости от того, какую информацию несут сигналы обратной связи (о степени напряжения мышц, положении частей тела и т.д.), они приходят в разные чувствительные центры мозга, и на разных уровнях переключаются на моторные пути. Он предположил, что для построения движений различной сложности «команды» отдаются на разных уровнях нервной системы. При автоматизации движений эта функция передается на более низкий уровень.
Бернштейн выделил следующие уровни:
1) А — самый низкий уровень. У человека он участвует в организации движений с другими уровнями, отвечает за тонус мышц, получает сигналы о степени напряжения мышц, информацию от органов равновесия. Он самостоятельно регулирует движения, связанные с вибрацией (дрожь), удержание позы в полетной фазе прыжка.
2) В — уровень синергий, отвечает за слаженные движения всего тела, ритмические и циклические движения, за автоматизацию двигательных навыков, выполняет внутреннюю координацию сложных движений, получает информацию о взаимном положении и движении частей тела. Движения этого уровня - потягивания, мимика.
3) C – уровень пространственного поля, обеспечивает ориентацию в пространстве, отвечает за движения, приспособленные к пространственным свойствам объектов (форме, положению, длине). Сюда поступает информация о внешнем пространстве. Движения: переместительные, баллистические.
4) D — уровень предметных действий. Это уровень коры головного мозга, отвечает за организацию действий с предметами в соответствии с их предметными назначениями. К нему относятся все орудийные действия и манипуляции с предметами. Здесь не фиксирован двигательный состав, движения могут быть разные, а конечный результат действия - одинаковый.
5) Е — уровень интеллектуальных двигательных актов. К нему относятся речевые движения, движения письма, символической, кодированной речи. Они определяются отвлеченным, вербальным смыслом, здесь речь идет о передаче знаний или замысла, что предполагает произвольный уровень регуляции действий. Бернштейн говорил, что этот уровень наименее изучен, и возможно, что здесь несколько уровней.
Чем сложнее, осмысленнее двигательная задача, тем более высок уровень построения движения, и тем более высокие уровни нервной системы участвуют в решении этой задачи и реализации движения.
Бернштейн Н.А. сделал важные выводы:
1) В организации сложных движений участвуют одновременно ведущий уровень, на котором строится движение, и все нижележащие (фоновые) уровни, но в сознании человека представлены только те компоненты движения, которые строятся на ведущем уровне, а работа фоновых уровней обычно не осознается.
2) Одно и то же движение может строиться на разных ведущих уровнях. Ведущий уровень построения движения определяется смыслом, или задачей, движения. Можно изменить характер протекания движения, изменив его смысл для исполнителя. Какие именно структуры участвуют в обеспечении построения движений человека, зависит от смысла движения. В подтверждение Бернштейн ссылался на исследование, проведенное во время войны: амплитуда движений раненой руки больного при решении двигательных задач менялась в зависимости от их смысла для испытуемого.
Данное положение показывает решающее значение такой психологической категории, как задача или цель движения, для организации и протекания физиологических процессов. Предполагается, это также свидетельствует о приоритете принципа активности над принципом реактивности.
27 Нейронные механизмы сочетания движения глаз, головы, рук.
Сравнительный анализ нервно-мышечной организации глазодвигательной активности и других соматических движений позволяет выявить черты как сходства, так и различия.
Моторика глаз подчинена главным принципам работы нервно-мышечной системы и то же время обладает определенной спецификой, обусловленной той ролью, которую выполняет двигательный аппарат глаза в процессе реализации зрительных функций.
Предельно высокая скорость глазных скачков способствует быстроте зрительных ориентировок (и оптомоторных реакций) и обеспечивает фильтрацию тех перемещений изображения по сетчатке, которые вызываются собственными движениями глаз. На пути решения этих задач произошла, по-видимому, дивергенция двух механизмов, которые в работе других мышц разделены в меньшей мере, а именно механизма рекрутирования двигательных единиц и механизма градуальных изменений частоты разрядов нейронов. В результате такой дивергенции иннервация саккад приобрела специфический "взрывной" характер, а плавные движения (при фовеальном прослеживании) управляются по принципу частотной регуляции мышечных сокращений. Проблема взаимодействия фазических и тонических единиц требует дальнейших фактических уточнений. Однако на сегодня она может быть гипотетически решена следующим образом. Наиболее вероятно, что "истинные" тонические единицы участвуют в эволюционно древних компенсаторных движениях глаз, афферентируемых вестибулярно.
В филогенезе нервная трубка начинает свертываться на уровне будущего продолговатого мозга, и уже отсюда происходит ее дальнейшее развитие в обе стороны. Это вполне объяснимо: именно здесь, в непосредственной близости от ротового отверстия, закладываются важнейшие вегетативные и соматические образования. Здесь формируется древнейший вегетативно-соматический "координационный центр", с которого и начинается процесс кефализации. В условиях примитивного червеобразного движения повороты переднего конца тела способствовали пространственной ориентации животного, возможности которой значительно расширились с появлением светочувствительных нервных окончаний - зачатков будущих глаз.
С этих позиций становится понятным смысл представительства моторного аппарата зрения в нижних оливах - парных ядрах продолговатого мозга, участвующих в регуляции поворотов головы и ориентации тела. Видимо, благодаря таким связям глазодвигательный аппарат современных позвоночных в ходе своего развития оказался включенным в интегральную схему пространственной ориентации животного, позволяющую координировать движения головы, глаз и всего тела. Поскольку ориентация и перемещение в пространстве неразрывно связаны с влиянием гравитации, уже на ранних этапах филогенеза позвоночных сформировался контур регулирования глазодвигательной активности, объединяющий вестибулярный аппарат, мозжечок и нижние оливы.
Глаза современных позвоночных сформировались в результате "выпячивания" нервной трубки на уровне среднего мозга. Поэтому исходную основу ретинального контура регулирования движений глаз составляют мезэнцефалические нервные образования. Верхние двухолмия (ВД), в которых оканчиваются аксоны ганглиозных клеток, - это, по существу, стволовые ганглии глазных сетчаток. По своему происхождению они могут рассматриваться как аналоги сегментарных афферентных ганглиев спинного мозга. В таком случае клетки глубоких слоев ВД и покрышки (тегментума) среднего мозга оказываются аналогами сегментарных интернейроов, роль которых заключается в мультисенсорной интеграции зрительных, слуховых и приприоцептивных (от мышц глаз, шеи и других) посылок. Полисенсорная конвергенция на нейронах ВД имеет, видимо, решающее значение для реализации двух механизмов:
а) соотнесения моторики глаз с общей соматической моторикой;
б) адекватных оптомоторных реакций, т.е. движений в ответ на зрительные сигналы. Нейроны глубоких слоев ВД посылают свои аксоны в парамедиальную ретикулярную формацию моста, а их коллатерали поступают непосредственно во все глазодвигательные ядра.
Таким образом, кратчайший путь, реализующий глазадвигательный ответ на зрительный стимул, включает в себя следующие инстанции: рецепторы сетчатки - ганглиозные клетки - клетки афферентного поверхностного слоя ВД - интернейроны ВД - премоторные нейроны глубоких слоев ВД - мотонейроны глазодвигательных ядер. ВД, по всей вероятности представляет собой тот уровень, на котором под управляющим воздействием идущих из коры (а также из других инстанций) импульсов, происходит селекция ретинальных сигналов, нужных для организации адекватной моторики в соответствии с текущими поведенческими задачами.
Говоря о роли ВД в регулировании глазодвигательной активности, нельзя не учитывать, что движения глаз могут афферентироваться другими сенсорными входами, не включающими ВД. Так для реализации вестибуло-окуломоторного рефлекса достаточно связей, охватывающих вестибулярных аппарат, ядра парамедиальной ретикулярной формации и глазодвигательные ядра. Нейроны ВД не активируются при плавных прослеживающих движениях глаз. Выдвинуто предположение, что в этом случае функционирует контур регулирования, содержащий в качестве центрального звена латеральные коленчатые тела. Таким образом, стволовой уровень глазодвигательного контроля представляет весьма сложную, относительно сепаратную (способную самостоятельно функционировать), филогенетически древнюю целостную систему, работающую в неразрывном единстве с сенсорными единицами этого же (стволового) уровня. Именно этим уровнем детерминируются основные динамические характеристики глазодвигательной активности, согласованная работа обоих глаз и координация движений глаз с другими моторными компонентами поведения. В этом случае неизбежно возникает вопрос о принципиальном значении и реальных механизмах высшего кортикального управления глазодвигательной активностью.
Активация фронтальных глазодвигательных полей связана не только с движениями глаз, но и с поворотами головы и играет, вероятно, важную роль в координации движений глаза и руки человека. Париетальная кора ответственна за точную пространственную организацию глазных движений в соответствии со "схемой тела" и изменениями положения тела в пространстве.
Таким образом, различные кортикальные зоны не просто "двигают" глаза, а организуют сложные синергии с участием движений глаз. Иерархический подход к организации движений глаз показывает, что вопросы о разделении функций "по вертикали", характер соподчинения отдельных этажей и их роль в инициации моторных компонентов поведения - сложная и далеко не решенная проблема. По-видимому, в реальных условиях поведения и деятельности взаимодействия внутри иерархии могут быть весьма вариативными. В разных ситуациях и при разных степенях обученности системы в целом ведущая роль может принадлежать разным звеньям и дихотомический выбор между двумя направлениями движения командных посылок (или моторных программ) - снизу вверх или сверху вниз - вовсе не является обязательным.
29 Сущность движения. Формирование навыка.
Психофизиологическое направление в психологии рассматривает движения человека в терминах рефлекторных актов, последовательно описываемых движение в виде комплекса рефлексов.
С позиций данного направления движение существует в форме двигательного комплекса (навыка). Понятие «навык» понимается как моторный акт, жестко «завязанный» с условиями его проведения. Рефлекс является единицей анализа двигательных, моторных актов человека. Для Вундта В. движение становится «бессознательным умозаключением», К.Д.Ушинский считает его «полурефлексом» (результатом воспитания), П.Ф.Лесгафт определяет человеческое движение «сознательной физической работой».
А.А.Ухтомский, исследуя физиологические свойства мышечного сокращения, выделил «живой активный хронотип», индивидуально характерный для каждого человека. В этом понятии А.А.Ухтомский характеризует человеческие свойства движений со стороны задействованного в движение времени и пространства индивидуального характера. Кроме того, Ухтомский А.А. отметил образование нервно-мышечного субстрата как состояния мышечной ткани, характерной для данного движения («нервно-мышечные констелляции»). Эти констелляции создаются неоднократным повторением одного и того же движения, они становятся дополнительной опорой в необходимом ограничении избыточных инерциальных проявлений движения. А.А.Ухтомский показал, что образование констелляций позволяет работающей мышце приобрести дополнительную экономичность ее работы в данном движении. Вот почему выученное новичком движение сразу не позволяет реализовать его возможности: констелляции должны «созреть».
Н.А.Бернштейн показал сложность человеческих действий и назвал их «живым! движениями». «Живое» наполнение человеческих движений Н.А.Бернштейн видел в том, что, кроме внешних воздействий на тело человека, человеку приходится учитывать и внутренние колебания конструкции тела. Еще более усложняется картина «живого движения» по Н.А.Бернштейну, когда человек выделяет цели и мотивы собственных движений в виде «образа потребного будущего», становящегося матрицей управления движениями. «В овладении движением Н.А.Бернштейн выделил три стадии: 1) стадия «излишнего закрепощения» движений, когда производится иммобилизация избыточных степеней свободы системы, 2) стадия уточнения степени участия в движении отдельных мышечных групп и 3) стадия точного использования всех сопутствующих движению сил инерции, тяжести, реактивных сил и энергозатрат». На примере биодинамики ходьбы и бега Н.А.Бернштейн показал, что «текущая коррекция» движений, как «живого» движения, представляет собой «повторение без повторения» и этим обеспечивает надежность воспроизведения движений человека и ее вариативность.
В исследованиях движений Н.А.Бернштейн доказал, что уровни управления физиологическими структурами движений выступают уровнями построения движения. В своей классификации структур построения движений Н.А.Бернштейн разделил их на:
1) познотонические (уровень «А»),
2) автоматизированные (уровень «В»),
3) сложные движения (уровень «С»),
4) «предметные действия» — действия с вещами (уровни «Д» и «Е»).
Такой подход давал возможность Н.А.Бернштейну утверждать, что разные стороны, свойства движения контролируются разными нервными структурами, своим специфическим способом и одновременно. Целеполагание всех движений контролируется сверху вниз. Высшим отделам мозга принадлежит и предвосхищение (антиципация) реализуемого движения, им строится, как считал Н.А.Бернштейн, «образ потребного будущего». Среднему мозгу Бернштейн отводил роль регулятора степени напряжения мышц, поддерживающего задаваемый темп и ритм движений, программирования последовательности включения и выключения задействованных мышц. Низшие отделы мозга принимают участие в каждом движении, обеспечивая определенный режим деятельности задействованием сосудистых, тонических, эндокринных и Других систем организма человека.
Последующие исследования движений человека реализованы в спортивной биомеханике: Д.Д.Донской, В.М.Зациорский, В.Б.Коренберг, В.Д.Мазннченко, А.Н.Мишин, В.Т.Назаров, В.Н.Тутевич и т.д. Все они опирались на результаты исследований Н.А.Бернштейна.
Наследие Н.А.Бернштейна разрабатывается и в трудах физиологов: А.С.Батуев, С.Г.Геллершгейн, В.С.Гурфинкель, Я.М.Коц, А.Н.Крестовников и др.
Заметное влияние на понимание проблем психологии движений оказали работы Б.Г.Ананьева. Он показал, что асимметрия восприятия левой и правой половины тела человека несет определенные функции в анализе пространственных представлений действующего человека. Он доказал, что одна стороне тела «специализируется» в сенсорном направлении обеспечения движений, другая — в реализации практических действий.
«В зарубежных исследованиях авторы описывают движение человека чисто прагматически. Чаще всего в анализе движений человека исследователи опираются на физиологическое знание рефлекторного акта или на его бихевиористическое понимание, где движения приравниваются к реакциям в системе «стимул — реакция». В этой диаде психологическое содержание несут стимулы, которые инициируют реакции. Анализ двигательной деятельности сводится к отысканий стимулов, которые реализуют соответствующие им реакции. Возникновение стимулов и содержание реакций обычно не рассматриваются, они как бы присутствуют изначально. Такой анализ не раскрывает содержания деятельности ни ее стороны стимула, ни со стороны реакции. В стимуле остается скрытым целеобразование и мотивация деятельности, а в реакциях за кадром у них остается пространственно-временная и практическая сторона деятельности. Этих взглядов на движение придерживаются, в основном, американские психологи, которые описывают эти проблемы в разрезе «моторного научения».
В своей докторской диссертации А.Леонтьев рассматривает генез деятельности, как животных, так и человека. Им последовательно представлено доказательство развития психики допсихической жизни, развитие сенсорной и перцептивной психики животных в соответствии с развитием их деятельности и восприятия. Эти данные имеют прямое отношение к анализу психического развития ребенка. В своей работе А.Н.Леонтъев выделяет развитие структурных компонентов деятельности человека, начиная с уровня перцептивной психики: деятельность, действие, операции, функциональный блок. Там же им дается психологическое определение комплексу движений, навыку, как автоматизированному образованию, исполняемому на более низком уровне в сравнении с уровнем своего происхождения.