Строение прокариотической клетки. Функции органелл.
Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации (рис.1). Структуры, расположенные снаружи от ЦПМ (клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки), называют обычно поверхностными структурами. Термином "клеточная оболочка" часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). ЦПМ вместе с цитоплазмой называется протопластом. Рассмотрим сначала строение, химический состав и функции поверхностных клеточных структур.

|
| Рис. 1. Комбинированное изображение прокариотной клетки. А — поверхностные клеточные структуры и внеклеточные образования: 1 — клеточная стенка; 2 — капсула; 3 — слизистые выделения; 4 — чехол; 5 — жгутики; 6 — ворсинки; Б — цитоплазматические клеточные структуры: 7 — ЦПМ; 8 — нуклеоид; 9 — рибосомы; 10 — цитоплазма; 11 — хроматофоры; 12 — хлоросомы; 13 — пластинчатые тилакоиды; 14 — фикобилисомы; 15 — трубчатые тилакоиды; 16 — мезосома; 17 — аэросомы (газовые вакуоли); 18 — ламеллярные структуры; В — запасные вещества: 19 — полисахаридные гранулы; 20 — гранулы поли-b-оксимасляной кислоты; 21 — гранулы полифосфата; 22 — цианофициновые гранулы; 23 — карбоксисомы (полиэдральные тела); 24 — включения серы; 25 — жировые капли; 26 — углеводородные гранулы (по Schlegel, 1972) |
Обязательные (основные органеллы)
Цитоплазматическая мембрана
Содержимое клетки отделяется от клеточной стенки цитоплазматической мембраной (ЦПМ) — обязательным структурным элементом любой клетки, нарушение целостности которого приводит к потере клеткой жизнеспособности. На долю ЦПМ приходится 8—15% сухого вещества клеток.
ЦПМ — белково-липидный комплекс, в котором белки составляют 50–75%, липиды — от 15 до 45%. Кроме того, в составе мембран обнаружено небольшое количество углеводов. Как правило, липиды и белки составляют 95% и больше вещества мембран. Главным липидным компонентом бактериальных мембран являются фосфолипиды — производные 3-фосфоглицерина. Хотя у прокариот найдено множество различных фосфолипидов, набор их в значительной степени родо- и даже видоспецифичен. Широко представлены в бактериальных мембранах различные гликолипиды. Стерины отсутствуют у подавляющего большинства прокариот, за исключением представителей группы микоплазм и некоторых бактерий. Так, в ЦПМ Acholeplasma содержится 10–30% холестерина, поглощаемого из внешней среды, от общего содержания мембранных липидов. Из других групп липидов в мембранах прокариот обнаружены каротиноиды, хиноны, углеводороды.
Структура мембран. Мембранные липиды всех эубактерий и части архебактерий образуют бислои, в которых гидрофильные "головы" молекул обращены наружу, а гидрофобные "хвосты" погружены в толщу мембраны (рис.2). Углеводородные цепи, прилегающие к гидрофильным "головам", довольно жестко фиксированы, а более удаленные части "хвостов" обладают достаточной гибкостью.

Рис.2Модель строения элементарной биологической мембраны: 1 — молекулы липидов: а — гидрофильная "голова"; б — гидрофобный "хвост"; 2 — молекулы белков: в — интегральная; г — периферическая; д — поверхностная.
Функции:
1. защитная
2. транспортная
3. участие в энергетическом обмене
4. участие в фотосинтезе
5. участие в распределении генетического материала
Функции 3-5 характерны только для прокариот
Функции ЦПМ прокариот. ЦПМ прокариот выполняет разнообразные функции, в основном обеспечиваемые локализованными в ней соответствующими ферментными белками. Первоначально была постулирована барьерная функция клеточной мембраны, получившая позднее экспериментальное подтверждение. С помощью специальных переносчиков, называемых транслоказами, через мембрану осуществляется избирательный перенос различных органических и неорганических молекул и ионов. В ней локализованы ферменты, катализирующие конечные этапы синтеза мембранных липидов, компонентов клеточной стенки и некоторых других веществ.
Общепризнана роль ЦПМ прокариот в превращениях клеточной энергии. У бактерий, источником энергии для которых служат процессы дыхания или фотосинтеза, в ЦПМ определенным образом расположены переносчики цепи электронного транспорта, функционирование которых приводит к генерированию электрохимической энергии (DmH+), используемой затем в клетке по разным каналам, в том числе и для образования химической энергии (АТФ). ЦПМ является одним из компонентов аппарата генерирования DmH+. В мембране расположены также ферментные комплексы, обеспечивающие превращения: DmH+ ® АТФ. ЦПМ принимает участие в репликации и последующем разделении хромосомы прокариотной клетки.
В последнее время выявляется еще одна функциональная грань клеточных мембран — их интегрирующая роль в организме, вполне сочетающаяся с давно установленной разъединяющей (барьерной) функцией. Клетка — единое целое. В обеспечении этого принципа клеточной организации важная роль принадлежит мембранам. Показан перенос электрохимической энергии и электронов вдоль мембран. Последние рассматриваются так же как возможные пути транспорта жирорастворимых субстратов и молекулярного кислорода.
ЦПМ является основным барьером, обеспечивающим избирательное поступление в клетку и выход из нее разнообразных веществ и ионов (У грамположительных форм ЦПМ является и единственным барьером такого рода, у грамотрицательных эубактерий функции дополнительного барьера (молекулярного "сита") выполняет наружная мембрана клеточной стенки, через которую молекулы транспортируются только по механизму пассивной диффузии.)Осуществляется это с использованием разных механизмов мембранного транспорта. Выделяют 4 типа транспортных систем, с участием которых происходит проникновение молекул в бактериальную клетку: пассивную диффузию, облегченную диффузию, активный транспорт и перенос химически модифицированных молекул.
Молекулы воды, некоторых газов (например O2, H2, N2) и углеводородов, концентрации которых во внешней среде выше, чем в клетке, проходят через ЦПМ внутрь клетки посредством пассивной диффузии. Движущей силой этого процесса служит градиент концентрации вещества по обе стороны мембраны. Основным соединением, поступающим в клетку и покидающим ее таким путем, является вода. Движение воды через мембрану, подчиняющееся законам пассивной диффузии, привело к выводу о существовании в мембране пор. Эти поры пока не удалось увидеть в электронный микроскоп, но некоторые данные о них были получены косвенными методами. Расчетным путем установлено, что поры должны быть очень мелкими и занимать небольшую часть поверхности ЦПМ. Высказывается предположение, что они не являются стабильными структурными образованиями, а возникают в результате временных перестроек молекулярной организации мембраны.
Большинство (если не все) гидрофильных веществ поступает в клетку за счет функционирования систем, в состав которых входят специальные переносчики (транслоказы, или пермеазы), так как скорость физической диффузии этих веществ через гидрофобный слой мембраны очень невелика. Переносчики — вещества белковой природы, локализованные в мембране и характеризующиеся высокой субстратной специфичностью,—связываясь с субстратом, подвергаются конформационным изменениям и вследствие этого приобретают способность к перемещению субстрата с одной стороны ЦПМ на другую.
Известен механизм транспорта, получивший название облегченной диффузии, требующий для переноса веществ через мембрану участия транслоказ. Перенос веществ в этом случае происходит по градиенту их концентрации и не требует энергетических затрат. Этот механизм транспорта не получил широкого распространения у прокариот. Основным механизмом избирательного переноса веществ через ЦПМ прокариот является активный транспорт, позволяющий "накачивать" в клетку молекулы и ионы против их концентрационных и электрических градиентов. Этот процесс, так же как и облегченная диффузия, протекает при участии локализованных в ЦПМ переносчиков белковой природы с высокой специфичностью к субстрату, но в отличие от облегченной диффузии для движения против электрохимического градиента требует затрат метаболической энергии. Транспорт такого рода должен быть поэтому сопряжен с реакциями, продуцирующими энергию в химической или электрохимической форме.
Во всех описанных выше путях переноса веществ через ЦПМ они поступают в клетку в химически неизмененном виде. У прокариот известны системы транспорта, с помощью которых осуществляется поступление в клетку ряда сахаров, при этом процесс их переноса через мембрану сопровождается химической модификацией молекул. Так происходит, например, поступление в клетки многих прокариот молекул глюкозы, в процессе которого они фосфорилируются.
Клеточная стенка
Клеточная стенка — важный и обязательный структурный элемент подавляющего большинства прокариотных клеток, располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с окружающей средой (у клеток, не содержащих этих слоев клеточной оболочки). На долю клеточной стенки приходится от 5 до 50% сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. Концентрация солей в клетке, как правило, намного выше, чем в окружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка чисто механически защищает клетку от проникновения в нее избытка воды.По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты, относящиеся к эубактериям, делятся на две большие группы. Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна: у так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются. У некоторых эубактерий положительная реакция при окрашивании описанным выше способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки.

|
| Рис. 3 Клеточная стенка грамположительных (А) и грамотрицательных (Б) эубактерий: 1 — цитоплазматическая мембрана; 2 — пептидогликан; 3 — периплазматическое пространство; 4 — наружная мембрана: 5 — цитоплазма, в центре которой расположена ДНК |
Клеточные стенки грамположительных и грамотрицательных эубактерий резко различаются как по химическому составу,так и по ультраструктуре (рис. 3).
В состав клеточной стенки эубактерий входят семь различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке. У грамположительны эубактерий он составляет основную массу вещества клеточной стенки (от 40 до 90%), у грамотрицательных — содержание пептидогликана значительно меньше (1—10%). Клеточная стенка цианобактерий, сходная с таковой грамотрицательных эубактерий, содержит от 20 до 50% этого гетерополимера.
Под электронным микроскопом клеточная стенка грамположительных эубактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм. У грамотрицательных эубактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2—3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8—10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных эубактерий получил название наружной мембраны.
бактерий плотно прилегает к ЦПМ, в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электронно-прозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической: и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп эубактерий, характерно только для грамотрицательных форм.
Функции:
1. защитная
2. придание формы клетки
3. антигенная
Цитоплазма
Содержимое клетки, окруженное ЦПМ, называется цитоплазмой. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: внутрицитоплазматическими мембранами (если они есть), генетическим аппаратом, рибосомами и включениями разной химической природы и функционального назначения.
Рибосомы — место синтеза белка — рибонуклеопротеиновые частицы размером 15—20 нм. Их количество в клетке зависит от интенсивности процессов белкового синтеза и колеблется от 5000 до 90 000. Общая масса рибосом может составлять примерно 1/4 клеточной массы, а количество рибосомальной РНК (рРНК) — 80–85% всей бактериальной РНК. Отношение рРНК/белок в рибосомах Е. coli составляет 2:1, у других прокариот оно может быть несколько сдвинуто в сторону преобладания белка. Рибосомы прокариот имеют константу седиментации 705, отчего получили название 70S-частиц. Они построены из двух неодинаковых субчастиц: 305- и 50S-субъединиц
Синтез белка осуществляется агрегатами, состоящими из рибосом, молекул информационной и транспортных РНК и называемыми полирибосомами, или полисомами. Последние могут находиться в цитоплазме или же быть связанными с мембранными структурами.
Генетический аппарат и репликация хромосомы
У прокариот ДНК представляет собой более или менее компактное образование, занимающее определенную область в цитоплазме и не отделенное от нее мембраной, как это имеет место у эукариот. Чтобы подчеркнуть структурные различия в генетическом аппарате прокариотных и эукариотных клеток, предложено у первых его называть нуклеоидом в отличие от ядра у вторых.
При электронно-микроскопическом наблюдении видно, что нуклеоид прокариот, несмотря на отсутствие ядерной мембраны, довольно четко отграничен от цитоплазмы, занимает в ней, как правило, центральную область и заполнен нитями ДНК диаметром около 2 нм.
Вся генетическая информация прокариот содержится в одной молекуле ДНК, имеющей форму ковалентно замкнутого кольца и получившей название бактериальной хромосомы(В прокариотной клетке ДНК может находиться и вне бактериальной хромосомы — в плазмидах, но последние не являются обязательными клеточными компонентами.)Длина молекулы в развернутом виде может составлять более 1 мм, т. е. почти в 1000 раз превышать длину бактериальной клетки. Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Однако если исходить из того, что молекула ДНК образует беспорядочный клубок, трудно объяснить процесс репликации и последующее распределение образовавшихся хромосом по дочерним клеткам. Специальные исследования показали, что хромосомы прокариот представляют собой высокоупорядоченную структуру, имеющую константу седиментации 1300—2000S для свободной и 3200—7000S для связанной с мембраной формы. В том и другом случае часть ДНК в этой структуре представлена системой из 20—100 независимо суперспирализованных петель. В обеспечении суперспирализованной организации хромосом участвуют молекулы РНК.
|
| Рис. 4 Репликация кольцевой бактериальной хромосомы в двух направлениях. А — родительская молекула ДНК; Б — промежуточные репликативные формы; В — дочерние молекулы ДНК после завершения процесса репликации и расхождения: 1 — точка начала репликации; черными стрелками показано направление репликации |
Хромосомы большинства прокариот имеют молекулярную массу в пределах 1—3x109 Да. В группе микоплазм генетический материал представлен молекулами, имеющими наименьшее для клеточных организмов количество ДНК (0,4–0,8x109), а наибольшее содержание ДНК обнаружено у нитчатых цианобактерий (8,5x109). Хотя каждая прокариотная клетка содержит 1 хромосому, часто в экспоненциально растущей культуре количество ДНК на клетку может достигать массы 3, 4, 8 и более хромосом. Нередко в клетках при действии на них определенных факторов (температуры, pH среды, ионизирующего излучения, солей тяжелых металлов, некоторых антибиотиков и др.) происходит образование множества копий хромосомы. При устранении воздействия этих факторов, а также после перехода в стационарную фазу в клетках, как правило, обнаруживается по одной копии хромосомы.
ДНК прокариот построена так же, как и эукариот. Молекула ДНК несет множество отрицательных зарядов, поскольку каждый фосфатный остаток содержит ионизированную гидроксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками — гистонами. В клетках подавляющего большинства прокариот не обнаружено гистонов, поэтому нейтрализация зарядов осуществляется взаимодействием ДНК с полиаминами (спермином и спермидином), а также с ионами Mg2+. В последнее время у некоторых архебактерий и цианобактерий обнаружены гистоны и гистоноподобные белки, связанные с ДНК. Содержание пар оснований А+Т и Г+Ц в молекуле ДНК является постоянным для данного вида организма и служит важным диагностическим признаком. У прокариот молярная доля ГЦ в ДНК колеблется в очень широких пределах: от 23 до 75%.

|
| Рис. 5 Механизм распределения бактериальных хромосом: А — бактериальная клетка содержит частично реплицированную хромосому, прикрепленную к мембране в точке (или точках) репликации; Б — репликация хромосомы завершена. В бактериальной клетке две дочерние хромосомы, каждая из которых прикреплена к ЦПМ. Показан синтез клеточной стенки и ЦПМ; В — продолжающийся синтез мембраны и клеточной стенки приводит к разделению дочерних хромосом. Показано начало деления клетки путем образования поперечной перегородки: 1 — ДНК; 2 — прикрепление хромосомы к ЦПМ: 3 — ЦПМ; 4 — клеточная стенка: 5 — синтезированный участок ЦПМ; 6 — новый материал клеточной стенки |
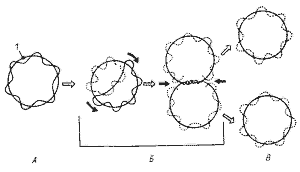
Деление молекулы ДНК (репликация) происходит по полуконсервативному механизму и в норме всегда предшествует делению клетки. С помощью электронного микроскопа установлено, что репликация ДНК начинается в точке прикрепления кольцевой хромосомы к ЦПМ, где локализован ферментативный аппарат, ответственный за репликацию. Часто можно обнаружить, что контакт ДНК с ЦПМ осуществляется посредством мезосом. Репликация, начавшаяся в точке прикрепления, идет затем в двух противоположных направлениях, образуя характерные для кольцевой хромосомы промежуточные структуры (рис.4). Возникающие дочерние хромосомы остаются прикрепленными к мембране. Репликация молекул ДНК происходит параллельно с синтезом мембраны в области контакта ДНК с ЦПМ. Это приводит к разделению (сегрегации) дочерних молекул ДНК и оформлению обособленных хромосом (рис.5)
Модель строения бактериальной хромосомы должна объяснять также прохождение в клетке процессов транскрипции и трансляции. Согласно существующим представлениям суперспирализованные петли соответствуют неактивным в данное время участкам ДНК и находятся в центре нуклеоида. По его периферии располагаются деспирализованные участки, на которых происходит синтез информационной РНК (иРНК), при этом, поскольку у бактерий процессы транскрипции и трансляции идут одновременно, одна и та же молекула иРНК может быть одновременно связана с ДНК и рибосомами (рис.6).

|
| Рис. 6. Модель организации нуклеоида Е. coli — наружная мембрана клеточной стенки; 2 — пептидогликановый слой; 3 — ЦПМ; 4 — точка прикрепления бактериальной хромосомы к ЦПМ; 5 — рибосомы, "сидящие" на иРНК. остальные объяснения см. в тексте (по Громову, 1985) |