1. Понятие о возбудимости и возбуждении. Показатели возбудимости.
2. Раздражители нервов и мышц.
3. Закон силы (силовых соотношений).
4. Закон "всё или ничего".
5. Действие постоянного тока на возбудимые ткани: а) закон полярного действия постоянного тока, б) электротон, периэлектротон.
6. Значение фактора времени в возникновении возбуждения.
7. Биоэлектрические явления в нервах и мышцах.
8. Лабильность, или функциональная подвижность тканей (Введенский)
9. Оптимум и пессимум частоты и силы раздражения (Введенский)
10.' Парабиоз, стадии его (Введенский)
11. Физиологические свойства нервных волокон и законы проведения возбуждения. по ним.
12.Аксоплазматический транспорт веществ и трофическая функция нервов
13. Механизм передачи возбуждения с нерва на мышцу (синаптическая передача возбуждения).
14. Физиологические свойства мышц.
15. Сила и работа мышц.
16. Понятие о механизме возникновения утомления.
К возбудимым тканям относятся нервная ткань (периферическая и ЦНС), мышцы (гладкие, скелетные, миокард) и железистые клетки. Возбудимость – это свойство (способность) тканей отвечать на раздражение. При этом ткань еще не находится в рабочем состоянии, а имеет только способность, готовность отвечать на раздражение. Возбуждение - это переход от состояния покоя к деятельности. Для возбуждения нервов характерна генерация потенциалов (импульса), а для мышцы - генерация биопотенциала и сокращение. По степени возбудимости ткани различаются. Наиболее высокая возбудимость у соматических нервов, но среди них есть волокна, обладающие неодинаковой возбудимостью и различной скоростью проведения возбуждения. Меньше, чем у соматических нервов, возбудимость вегетативной нервной системы (симпатической и парасимпатической). У мышц наибольшей возбудимостью обладают скелетные (которые сокращаются фазно, быстро - это главным образом мышцы конечностей). Меньшая возбудимость у тонических мышц (поддерживают позу, положение в пространстве), чем у фазных. Ещё меньше возбудимость у миокарда (у него очень большая абсолютная рефрактерность, занимающая всю систолу); самая маленькая возбудимость у гладких мышц (они сокращаются по принципу тонического сокращения).
Показатели возбудимости: I) порог раздражения – это минимальная сила раздражителя, вызывающая минимальную ответную реакцию (возбуждение). При высокой возбудимости тканей порог ниже, и наоборот. Подпороговое раздражение (сила раздражения ниже пороговой величины) - обычно не вызывает видимых изменений, но может привести к местному возбуждению без распространения на другие участки. Надпороговое раздражение - величина раздражителя выше пороговой величины, поэтому ответная реакция больше, и может быть максимальной на эти раздражения. 2) хронаксия - это минимальное время, которое необходимо для возникновения минимального возбуждения при силе тока в два порога (2 реобазы; реобаза – это иначе порог раздражения). Виды хронаксии: а) двигательная - критерием является сокращение мышцы. Для определения хронаксии какой-либо мышцы есть специальные таблицы с указанием расположения двигательных точек, по которым определяют место вхождения двигательного нервного окончания в какую-либо мышцу. При раздражении этой точки (двигательная точка) можно получить изолированное сокращение какой - либо мышцы. Двигательная хронаксия является объективным методом исследования (от воли человека ничего не зависит). Чем выше возбудимость мышц, тем ниже хронаксия. Например, хронаксия мышц сгибателей у человека ниже хронаксии мышц разгибателей, т.е. у первых возбудимость больше.
б) чувствительная хронаксия определяется по минимальному ощущению прохождения тока. Метод является субъективным. Характеризует состояние проводимости и рецепторного аппарата, в) рефлекторная хронаксия - раздражаются рецепторы на коже и в ответ на раздражение рецепторов возникает моторный ответ. Раздражение распространяется по рефлекторному пути (рецептор – афферентный путь – центр – эфферентный путь - мышца), г) субординационная - величина хронаксии соматических нервов может быть изменена под влиянием ЦНС. Если будет торможение ЦНС, то хронаксия может увеличиваться, при высокой возбудимости ЦНС все виды хронаксии снижены, д) конституциональная - когда исключается влияние ЦНС (например, в результате травмы может быть полный перерыв нерва). В начальном периоде после прекращения влияния ЦНС, хронаксия удлиняется, но потом может быть снижение, или восстановление до нормы. 3) лабильность (функциональная подвижность) - это скорость протекания каждого цикла возбуждения. При высокой возбудимости - лабильность выше, и наоборот.
Раздражители мыши и нервов. Все раздражители бывают 2-х групп: I) адекватные (естественные), например, нервный импульс адекватный раздражитель для мышц, ЦНС и нервов. 2) неадекватные (неестественные) - воздействие электрическим током, химическими веществами, механические воздействия, температура, дозированный электрический ток. Большое значение имеет вид тока. Чаще используется постоянный прямоугольный ток, так как раздражение постоянным током проявляется или в момент замыкания, или размыкания (резкого изменения величины тока). В период прохождения постоянного тока мышца не сокращается. Если нарастание тока будет постепенное, то ток может быть уже надпороговым, а сокращения мышцы не будет. Кроме крутизны (спада и нарастания), учитываются еще: I) величина (амплитуда) тока, 2) частота (Гц) - если она будет очень высокой, то большая часть раздражений будет попадать на абсолютную рефрактерность, и будут не эффективны. 3) продолжительность действия каждого стимула (в миллисекундах). Поэтому дозированный ток находит широкое применение. Другие виды неадекватных воздействий не нашли применения (так как их трудно дозировать). Хотя в естественных условиях такие раздражители, как химические в организме представлены широко (гормоны, медиаторы, другие биологически активные вещества).
Зависимость между величиной раздражителя и ответной реакцией: I) закон силы - с увеличением силы раздражения ответная реакция возрастает, но только до определённого предела. При какой-либо большой силе, может быть снижение ответной реакции. Этот закон характерен для всех возбудимых тканей. 2) закон "всё или ничего" - если величина раздражителя достигает пороговой, то может быть только полная реакция, а если величина эта низкая, ниже порога, то ничего нет. Однако если рассмотреть приложение этого закона к целой мышце или нервному стволу, а не к отдельному нервному или мышечному волокну, то этот закон не применим, так как в составе нервного ствола или мышцы имеются волокна, обладающие различной возбудимостью. Поэтому одни мышечные или нервные волокна будут реагировать на меньшие силы раздражителей, а другие - на большие. Поэтому с увеличением силы раздражителя, постепенно увеличивается сила сокращения скелетных мышц.
Действие постоянного тока на возбудимые ткани. Для постоянного тока (гальванической возбудимости тканей) характерны законы: I) закон полярного действия: а) постоянный ток действует своими полюсами - катодом (К) и анодом (А), б) в момент замыкания раздражающее действие оказывает катод, а в момент размыкания – анод, в) раздражающее действие катода сильнее, чем анода, поэтому порог для катода будет меньше, чем анода. Закон можно обнаружить на нервно-мышечном препарате. При слабом токе проявляется только замыкательное сокращение (ЗС); если ток средней силы, то возникает и замыкательное сокращение и размыкательное сокращение (PC). Если использовать сильный постоянный ток, то ответная реакция на раздражение зависит от расположения электродов, т.е. от направления тока. Если ближе к мышце располагается анод, то говорят, что ток восходящий. При расположении электродов в обратном порядке (ближе к мышце располагается катод) – ток нисходящий. При действии сильного тока под анадом происходит блокада проведения возбуждения (в этом месте происходит гиперполяризация), поэтому, возбуждение, возникшее под катодом дойдёт до анода, но до мышцы через участок гипрполяризации не пройдёт, и катодно-замыкателъного сокращения не будет.
Действие тока. средней силы. При любом расположении электродов будет и замыкательное и размыкательное сокращение. Если перевязать нерв между электродами, то в зависимости от расположения электродов будет:
а) если электроды расположены: ближе к мышце находится катод, а за участком перевязки – анод, то замыкательное сокращение есть, а размыкательного - нет, так как возбуждение под анодом дойдя до перевязки, дальше не распространяется, и мышца не сокращается.
б) при расположении электродов: ближе к мышце находится анод, а за участком перевязки нерва – катод, то при замыкании, импульсы не доходят до мышцы, и замыкательного сокращения не происходит.
Для определения электродиагностической формулы (в медицине), основанной на законе полярного действия постоянного тока, используют униполярный метод раздражения. Один электрод в виде пластинки накладывают на определённый участок тела, а другой - точечный электрод - на двигательную точку. Для униполярного метода характерно то, что раздражающим свойством обладает электрод с малой поверхностью (активный электрод), а пластинчатый - пассивный, нераздражающий электрод, так как плотность тока на единицу площади в точечном электроде во много раз больше, чем у пластинчатого. При наложении этих электродов будет 4 электрода: I) истинный катод (К), 2) истинный анод (А) 3) силовые линии идут от анода к катоду, которые пересекают нерв, входя в него. Затем они выходят из нерва, образуя дополнительный полюс - физиологический катод (К). 4) Затем силовые линии входят в нерв, и под катодом образуется физиологический анод (А). Если используется слабый ток, то определяется только катодно-замыкательное сокращение (КЗС). При токе
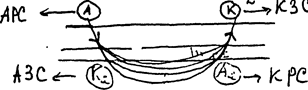
средней силы, определяется КЗС, анодно – замыкательное (АЗС) и анодно-размыкательное сокращение (АРС). При сильном токе можно определить все пороги (порог КЗС<АЗС<АРС<КРС). Это. и есть электродиагностическая формула. Для того чтобы понять кокой порог за счет какого электрода определяется, необходимо вспомнить закон полярного действия постоянного тока. Исходя из этого закона можем констатировать: КЗС - соответствует закону полярного действия, и сокращение мышцы происходит за счет раздражающего действия истинного катода; АЗС - не соответствует этому закону полярного действия, но в данном случае раздражающим электродом является физиологический катод (К²); АРС - соответствует закону полярного действия, раздражение происходит за счёт истинного А; КРС - не соответствует закону, но под катодом образуется физиологический А и за счёт раздражения этого полюса происходит КРС. Электродиагностическая формула определяется для диагностики нарушений целостности нерва, иннервирующего мышцу, и для контроля за ходом лечения. Например, при травме нерва, если происходит ущемление или нарушение целостности нерва, то электродиагностическая формула изменяется.
Электротон - это изменение возбудимости и проводимости под электродами постоянного тока. При замыкании или пропускании постоянного тока возбудимость под катодом увеличивается - это катэлектротон. В то же самое время, под анодом возбудимость и проводимость снижены - это анэлектротон. Анодом можно достичь полной блокады проведения нервного импульса. При продолжительном действии постоянного тока, или пропускании сильного постоянного тока, могут быть извращения обычных электротонических изменений: I) катодическая депрессия (описал Вериго) - при пропускании сильного постоянного тока, или длительном пропускании постоянного тока, снижается возбудимость и проводимость под катодом. 2) анодическое облегчение - пропускание сильного тока, или длительное пропускание тока приводит к повышению возбудимости под анодом. По принципу злектротонического изменения может быть проведение возбуждения в некоторых безмиелиновых волокнах, но скорость его ниже, чем импульсного. Кроме электротонических изменений, есть периэлектротонические изменения, имеющие противоположный характер, по сравнению с электротоническими: около анода возбудимость повышена, а около катода - возбудимость и проводимость снижены (явление периэлектротона описал Н.Е.Введенский).
Значение фактора времени для раздражения. Постоянный ток раздражающим действием обладает в момент замыкания и размыкания, если использовать длительные стимулы (в виде замыкания и размыания без учета продолжительности каждого стимула). Когда используются длительные интервалы раздражения, то время не имеет значения, а имеет значение лишь быстрота изменения величины тока в момент замыкания и размыкания (закон Дюбуа –Реймона). Но когда интервалы раздражения краткие (миллисекунды), то фактор времени имеет важное значение в возникновении возбуждения. Продолжительность времени раздражения и величина порога друг от друга зависят: с уменьшением времени раздражения, пороговая величина тока увеличивается (это видно на кривой Гоорвега – Вейса). Французские учёные Лапик и Бургиньон предложили определять не всю кривую, а только в точке хронаксии, которая определяется при удвоенной реобазе. Поэтому методика определения возбудимых тканей облегчается. Интервал с момента появления зависимости между временем и величиной порогового тока будет слева от ДС, а справа от этого - бесконечное время - здесь время не имеет значения для возникновения раздражения. Здесь действует закон Дюбуа-Реймона, который считал, что раздражающее действие постоянного тока зависит от быстроты изменения величины тока: в момент замыкания крутизна тока растёт, и в момент размыкания - она быстро падает. Продолжительность тока не оказывает раздражающего действия, здесь ток равен
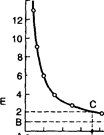
А D 2.0 мсек
. Кривая "сила — длительность" (кривая Гоорвега – Вейса) АВ — реобаза; 2 — двойная реобаза;AD — хронаксия. По оси абсцисс — продолжительность действия стимула, по оси ординат — величина реобазы.
пороговой величине (реобазе). В левой части кривой от ДC - действует зависимость между временем раздражения и величиной порогового тока (с уменьшением времени, пороговая величина тока увеличивается). Ток ультравысокой частоты (УВЧ) не обладает раздражающим действием, так как каждое последующее раздражение падает на абсолютную рефрактерность.
Биоэлектрические явления в нервах и мышцах. В 1791 г. Гальвани открыл. биотоки. Он работал с нервно – мышечными препаратами, их развешивал на медную проволоку у себя на балконе, а перила балкона были металлические и однажды, когда появился ветер, препараты (лапки лягушки) задевали металлические перила. Каждое соприкосновение с перилами сопровождалось сокращением лапок. Он сделал вывод, что за счет «животного электричества» (позднее названными биотоками) происходит сокращение лапок лягушки. Уже на следующий год физик Вольта выступил с критикой положений Гальвани. Вольта считал, что в данном случае речь идет о возникновении электродвижущей силы между разнородными металлами (в этом отношении он был прав), что и является раздражителем для нервно-мышечного препарата. В доказательство своей правоты Гальвани поставил опыт без использования металла. Он брал два нервно – мышечных препарата, нерв первого он раздражал, а нерв второго – накладывал на мышцу первого препарата (опыт Маттеучи).
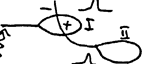
Раздражение нерва первого препарата всегда сопровождалось сокращением мышцы у первого и у второго препаратов. В момент возникновения возбуждения в мышце первого препарата, там появляется разность потенциалов между участками мышцы. Так как поверхность участка возбуждения заряжается электроотрицательно, а невозбужденный участок имеет положительный заряд, то эта разность потенциалов является раздражителем для мышцы второго препарата. Эти открытия окончательное подтверждение нашли в опыте Маттеучи «со вторичным тетанусом», когда он вызывался биотоками возбужденной мышцы другого препарата. В 1902 г. была сформулирована первая гипотеза Бернштейна о происхождении биотоков - мембранно-ионная гипотеза возникновения возбуждения. Эта теория существовала до 40-х годов 20 века, когда появилась возможность использования усилителей.
Мембранный потенциал (потенциал покоя)
Мембрана любой клетки состоит из липидов и белков (paньше думали, что имеются поры, через которые проходят электролиты внутрь и из клетки). Оказалось, что эти поры имеют функциональный характер (открываются в определённый момент). Любая живая клетка обладает способностью создавать концентрационный градиент: в цитоплазме концентрация ионов К+ ~в 50 раз больше, чем вне клетки, а натрия в 10 раз больше вне клетки, чем внутри её.
-70 мВ А
Опыт Ходжкина—Хаксли на гигантском аксоне кальмара; А — форма потенциала, зарегистрированная в опыте..
На схеме опыта Ходжкина – Хаксли показан скачок отрицательного потенциала в момент введения электрода внутрь аксона, т. е. внутренняя среда аксона была заряжена отрицательно относительно внешней среды.
Электрический потенциал содержимого живых клеток принято измерять относительно потенциала внешней среды, который обычно принимают равным нулю. Поэтому считают синонимами такие понятия, как трансмембранная разность потенциалов в покое, потенциал покоя, мембранный потенциал. Обычно величина потенциала покоя колеблется от —70 до —95 мВ. Величина потенциала покоя зависит от ряда факторов, в частности от избирательной проницаемости клеточной мембраны для различных ионов; различной концентрации ионов цитоплазмы клетки и ионов окружающей среды (ионной асимметрии); работы механизмов активного транспорта ионов. Все эти факторы тесно связаны между собой, и их разделение имеет определенную условность.
В невозбужденном состоянии клеточная мембрана высокопроницаема для ионов калия и малопроницаема для ионов натрия. Это было показано в опытах с использованием изотопов натрия и калия: спустя некоторое время после введения внутрь аксона радиоактивного калия его обнаруживали во внешней среде. Таким образом, происходит пассивный (по градиенту концентраций) выход ионов калия из аксона. Добавление радиоактивного натрия во внешнюю среду приводило к незначительному повышению его концентрации внутри аксона. Пассивный вход натрия внутрь аксона несколько уменьшает величину потенциала покоя.
Разность концентраций ионов калия вне и внутри клетки и высокая проницаемость клеточной мембраны для ионов калия обеспечивают диффузионный ток этих ионов из клетки наружу и накопление избытка положительных ионов К+ на наружной стороне клеточной мембраны, что противодействует дальнейшему выходу ионов К из клетки. Диффузионный ток ионов калия существует до тех пор, пока стремление их двигаться по концентрационному градиенту не уравновесится разностью потенциалов на мембране. Эта разность потенциалов называется калиевым равновесным потенциалом.
Равновесный потенциал (для соответствующего иона) — разность потенциалов между внутренней средой клетки и внеклеточной жидкостью, при которой вход и выход иона уравновешен (химическая разность потенциалов равна электрической). Следует иметь в виду: 1) состояние равновесия наступает в результате диффузии лишь очень небольшого количества ионов (по сравнению с их общим содержанием); калиевый равновесный потенциал всегда больше (по абсолютному значению) реального потенциала покоя, поскольку мембрана в покое не является идеальным изолятором, в частности имеется небольшая утечка ионов Na+.
В состоянии покоя клеточная мембрана высокопроницаема не только для ионов К+. У мышечных волокон мембрана высокопроницаема для ионов Сl. В клетках с высокой проницаемостью для ионов хлора, как правило, оба иона (Сl и К+) практически в одинаковой степени участвуют в создании потенциала покоя.
Известно, что в любой точке электролита количество анионов всегда соответствует количеству катионов (принцип электронейтральности), поэтому внутренняя среда клетки в любой точке электронейтральна. Действительно, в опытах Ходжкина, Хаксли и Катца перемещение электрода внутри аксона не выявило различие в величине потенциала покоя. Поддерживать постоянную разность концентрации ионов (ионную асимметрию) без специальных механизмов невозможно. В мембранах существуют системы активного транспорта, работающие с затратой энергии и перемещающие ионы против градиента концентраций. Экспериментальным доказательством существования механизмов активного транспорта служат результаты опытов, в которых активность АТФазы подавляли различными способами, например сердечным гликозидом оуабаином. При этом происходило выравнивание концентраций ионов К+ вне и внутри клетки и мембранный потенциал уменьшался до нуля.
Важнейшим механизмом, поддерживающим низкую внутриклеточную концентрацию ионов Na+ и высокую концентрацию ионов К+ является натрий - калиевый насос. Известно, что в клеточной мембране имеется система переносчиков, каждый из которых связывается с 3 находящимися внутри клетки ионами Na+ и выводит их наружу. С наружной стороны переносчик связывается с 2 находящимися вне клетки ионами К+, которые переносятся в цитоплазму. Работа систем переносчиков обеспечивается за счет АТФ. В результате чего обеспечивается: сохранение высокой концентрации К+ и низкой концентрации Nа + внутри клетки; калий – натриевый насос способствует сопряженному транспорту аминокислот и сахаров через клеточную мембрану. Потенциал действия
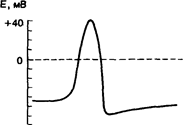
Под потенциалом действия понимают быстрое колебание потенциала покоя, сопровождающееся, как правило, перезарядкой мембраны. Потенциал действия появляется при нанесении раздражения. На кривых при регистрации потенциала действия регистрируются: I) латентный период (скрытый). 2) фаза деполяризации – крутой подъем кривой, при этом поверхность клетки заряжается отрицательно. 3) фаза реполяризации - восстановление прежнего состояния. Восстановления до исходного уровня сразу не происходит, а есть 4) следовые потенциалы (отрицательный и положительный).
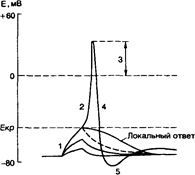
Потенциал действия одиночной клетки и его фазы. Реакция клеточной мембраны на раздражающий стимул; I — локальный ответ; 2 — быстрая деполяризация; 3 — реверсия; 4 — реполяризация; 5 —следовые (отрицательный и положительный) потенциалы.
. Активные подпороговые изменения мембранного потенциала называются локальным ответом.
Смещение мембранного потенциала до критического уровня приводит к генерации потенциала действия. Минимальное значение тока, необходимого для достижения критического потенциала, называют пороговым током. В опытах Ходжкина и Хаксли был обнаружен, на первый взгляд, удивительный эффект. Во время генерации потенциала действия мембранный потенциал уменьшался не просто до нуля, как следовало бы из уравнения Нернста, но изменил свой знак на противоположный. Анализ ионной природы потенциала действия, проведенный первоначально Ходжкиным, Хаксли и Катцем, позволил установить, что фаза деполяризации потенциала действия и перезарядка мембраны (овершут) обусловлены движением ионов натрия внутрь клетки, т. е. натриевые каналы оказались электроуправляемыми. Возбуждение приводит к активации натриевых каналов и увеличению натриевого тока. Это обеспечивает локальный ответ. Смещение мембранного потенциала до критического уровня приводит к стремительной деполяризации клеточной мембраны и обеспечивает фронт нарастания потенциала действия. Если удалить ионы Na из внешней среды, то потенциал действия не возникает. Аналогичный эффект удавалось получить при добавлении в перфузионный раствор специфического блокатора натриевых каналов – тетродоксина. При замене ионов натрия на другие ионы и вещества, например холин, удалось показать, что входящий ток обеспечивается натриевым током, т. е. в ответ на деполяризующий стимул происходит повышение натриевой проводимости. Таким образом, развитие фазы деполяризации потенциала действия обусловлено повышением натриевой проводимости.
Перезарядка мембраны, или овершут, весьма характерна для большинства возбудимых клеток. Амплитуда овершута характеризует состояние мембраны и зависит от состава вне- и внутриклеточной среды. На высоте овершута потенциал действия приближается к равновесному натриевому потенциалу, поэтому происходит изменение знака заряда на мембране. Экспериментально было показано, что амплитуда потенциала действия практически не зависит от силы стимула, если он превышает пороговую величину. Поэтому принято говорить, что потенциал действия подчиняется закону "все или ничего".
На пике потенциала действия проводимость мембраны для ионов натрия начинает быстро снижаться. Этот процесс называется инактивацией. Скорость и степень натриевой инактивации зависят от величины мембранного потенциала, т. е. они потенциалзависимы. При постепенном уменьшении мембранного потенциала до —50 мВ (например, при дефиците кислорода, действии некоторых лекарственных веществ) система натриевых каналов полностью инактивируется и клетка становится невозбудимой.
Потенциалзависимость активации и инактивации в большой степени обусловлена концентрацией ионов кальция. При повышении концентрации кальция значение порогового потенциала увеличивается, при понижении — уменьшается и приближается к потенциалу покоя. При этом в первом случае возбудимость уменьшается, во втором — увеличивается.
В обычных условиях задержанный выходящий калиевый ток существует некоторое время после генерации потенциала действия и это обеспечивает гиперполяризацию клеточной мембраны, т. е. положительный следовой потенциал. Положительный следовой потенциал может возникать и как следствие работы натриево-электрогенного насоса..
Инактивация натриевой системы в процессе генерации потенциала действия приводит к тому, что клетка в этот период не может быть повторно возбуждена, т. е. наблюдается состояние абсолютной рефрактерности.
Постепенное восстановление потенциала покоя в процессе реполяризации дает возможность вызвать повторный потенциал действия, но для этого требуется сверхпороговый стимул, так как клетка находится в состоянии относительной рефрактерности.
Исследование возбудимости клетки во время локального ответа или во время отрицательного следового потенциала показало, что генерация потенциала действия возможна при действии стимула ниже порогового значения. Это состояние супернормальности, или экзальтации.
В условиях покоя разность между наружной поверхностью мембраны и цитоплазмой существует постоянно. Если предварительно извлечь цитоплазму клетки, и внутрь клетки ввести раствор, с повышенным содержанием ионов натрия, то величина потенциала резко изменится. Поэтому решающее значение в возникновении потенциала покоя имеют ионы калия и натрия. Все электролиты имеют гидратную оболочку, но гидратная оболочка для ионов калия меньше, чем натрия. Поэтому ионы натрия не могут проходить в покое через мембрану. В образовании мембранного потенциала кроме этих ионов, принимают участие ионы хлора, расположенные под мембраной, и ионы кальция. Решающим фактором в возникновении заряда является наличие белковых молекул.
В гиперкалиевом растворе потенциал действия существенно снижается. В гипернатриевом растворе его величина увеличивается. Для анализа потенциала действия используются также фармакологические вещества - они обладают способностью блокировать или калиевый или натриевый канал. При блоке натриевого канала потенциал действия снижается. Это очень важно в диагностике инфаркта миокарда, опухоли головного мозга и др. В зависимости от того, как электроды расположены к различным здоровым и больным участкам, можно зарегистрировать двухфазный или однофазный потенциал действия.
Двухфазный потенциал действия регистрируется в том случае, если электроды находятся на здоровом – неповрежденном участке ткани. Если к двум точка приложить отводящие электроды, а к другой точке (показано стрелкой) - раздражающие электроды, то при нанесении искусственного раздражения будет двухфазное колебание потенциала. Отводящие электроды соединены с регистрирующей аппаратурой. Механизм возникновения двухфазного потенциала действия заключается в том, что наружная поверхность клетки, мышечного или нервного волокна имеет положительный заряд, а цитоплазма – отрицательный.
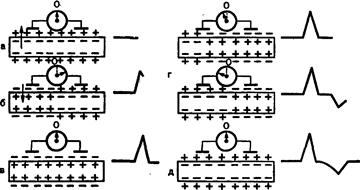
Динамика электрических потенциалов в мышечном волокне
Поэтому при регистрации потенциала вначале будет просто прямая линия (а), б) волна возбуждения проходит через участок под первым электродом. Наружная поверхность мембраны в этом участке становится отрицательной и между электродами возникает разность потенциалов, стрелка отклоняется, кривая поднимается вверх, в) затем возбуждение занимает всю поверхность между электродами, разность потенциалов исчезает и стрелка возвращается обратно в начальное состояние, кривая идёт вниз. г) под первым электродом происходит реполяризация (положительный заряд), а под вторым электродом имеет место еще деполяризация и стрелка гальвонометра отклоняется уже в другую сторону, и кривая идёт вниз. д) возбуждение покидает пределы второго электрода, под ним происходит реполяризация, и стрелка гальванометра возвращается в начальное положение.
Если один из электродов расположен на повреждённом участке, то стрелка не занимает нулевого положения, так как здоровый участок положительный, а повреждённый - отрицательный, и стрелка будет заранее отклонена. При таком расположении электродов регистрируется однофазный потенциал действия.
Это имеет важное значение для диагностики инфаркта миокарда, т.к. участок повреждения за ранее будет иметь отрицательный поверхностный заряд и волна возбуждения, которая будет распространяться на своем пути встретит измененный участок, а, следовательно, форма ЭКГ будет изменена.
Изменение возбудимости тканей при возбуждении.
Все возбудимые ткани при возбуждении меняют свою возбудимость, нервы сразу после нанесения раздражения, т. е. имеют очень небольшой латентный период. На рисунке представлены: наверху потенциал действия, внизу - изменение возбудимости нервного волокна в разные периоды возбуждения (абсолютная рефрактерность соответствует пику высоковольтного потенциала, относительная рефрактерность – фазе реполяризации, супернормальный период – отрицательному следовому потенциалу). Ниже в тексте приведена последовательность развития разных фаз:

местный Положительный следовой потенциал
процесс
Супернормальный период
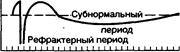
Время, мс
местный процесс, фаза деполяризации, фаза реполяризацпи, отрицательный следовый потенциал, положительный следовый потенциал, а также фазы изменения возбудимости нервного волокна: фаза абсолютной рефрактерности, фаза относительной рефрактерности, супернормальная возбудимость, субнормальная возбудимость и исходный уровень возбудимости. В фазу абсолютной рефрактерности возбудимость падает до нуля. Это соответствует фазе деполяризации. Максимальная рефрактерность наблюдается в момент пика деполяризации. Если стойкая деполяризация будет вызвана каким-либо веществом, то ткань теряет возможность отвечать на приходящее следующее возбуждение. Практически, торможение может быть при стойкой гиперполяризации, при стойкой деполяризации и стойкой поляризации, когда под действием какой-либо причины поверхностный положительный заряд не меняется.
Периоду реполяризации соответствует фаза относительной рефрактерности. Здесь возбудимость постепенно восстанавливается. После относительной рефрактерности, наступает фаза супернормальной возбудимости - соответствует отрицательному следовому потенциалу, затем наступает субнормальная возбудимость - соответствует положительному следовому потенциалу, и затем возбудимость приходит к исходному уровню
Кривые одиночного сокращения (I) и изменения возбудимости (2) скелетной, сердечной и гладкой мышц.
: 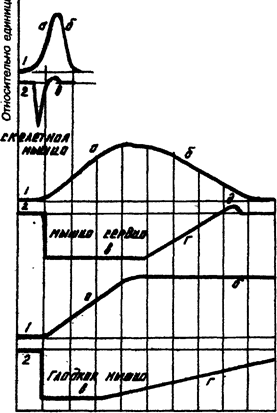
время, 0,1 с
а) период сокращения, б) период расслабления, в) фаза абсолютной рефрактерности, г) фаза относительной рефрактерности, д) фаза экзальтации (супернормальной возбудимости).
Разные мышцы обладают разной рефрактерностью и это свойство в значительной степени определяет особенности сократительной функции этих мышц.
Если взять постоянную частоту раздражающего тока, но постепенно увеличивать силу раздражения, то обнаружится, что с увеличением силы раздражения, ответная реакция будет увеличиваться. Такая же закономерность наблюдается в том случае, если увеличивать частоту наносимых раздражений при постоянной силе тока. Однако увеличение сокращения будет происходить до какой – оптимальной силы или частоты наносимых раздражений. Для оценки способности возбудимой ткани отвечать на раздражения разной частоты было введено понятие «лабильность» или функциональная подвижность (Н. Е. Введенский).
Под лабильностью понимают скорость протекания каждого цикла возбуждения или способность тканей воспроизводить без искажений частоту наносимых раздражений (для нервов лабильность ~ 1000 Гц, для скелетных мышц - ~250-500 Гц, для синапсов - ~ 100 Гц). Если частота наносимых раздражений больше лабильности, то не все импульсы будут воспроизводиться, а только те, которые не превышают величину лабильности (например, если на нерв нанести раздражения с частотой 2000 Гц, то ответов получим только 1000). При дальнейшем увеличении частоты, ответная реакция может исчезнуть. Для объяснения этого явления необходимо прибегнуть к понятиям абсолютной и относительной рефрактерности. Часть высокочастотных раздражений попадает на абсолютную рефрактерность, поэтому они ответа не вызывают. На основе лабильности Введенский разработал понятия об оптимуме и пессимуме силы и частоты раздражения. Та частота, при которой получается максимальный ответ - это оптимум частоты. Снижение ответной реакции, в связи с дальнейшим увеличением частоты наносимых раздражений называется пессимумом. Пессимум выражен тем больше, чем больше частота. Например, изменение возбудимости при одиночном сокращении скелетной мышцы:
При частоте больше 50 Гц возникает тетаничсское сокращение. Если наносить раздражение через интервалы одного цикла, то получится каждый раз одиночное сокращение. Если частоту увеличивать, интервалы между наносимыми раздражениями уменьшатся, и сокращение будет вначале ввиде зубчатого, а затем при дальнейшем увеличении частоты появится гладкий (сплошной) тетанус. Оптимум сокращения будет соответствовать фазе пика экзальтации - самой высокой возбудимости. При этом сила тока будет одна и таже, но так как возбудимость мышцы больше, то ответная реакция будет максимальной. При дальнейшем увеличении частоты, временные интервалы смешаются в фазу относительной рефрактерности, и часть импульсов при пессимуме попадает на эту фазу. Здесь возбуждение ниже, чем в фазу экзальтации, и ответная реакция будет ниже. Дальнейшее увеличение частоты приводит к попаданию импульсов на абсолютную рефрактерность. При этом нет ответа, т. к. в этот период полностью отсутствует возбудимость. Поэтому с лечебной целью используют токи высокой частоты, электроды прикладывают к коже, но мышцы не реагируют (не сокращаются), т. к. лабильность мышц намного ниже, чем частота тока УВЧ, и каждый стимул попадает на период абсолютной рефрактерности. Когда идёт множество импульсов из ЦНС по нерву к мышцам, то в зависимос<