Гликолиз
Глико́лиз (фосфотриозный путь, или шунт Эмбдена — Мейерхофа) — ферментативный процесс последовательного расщепления глюкозы в клетках, сопровождающийся синтезом АТФ. Гликолиз при аэробных условиях ведёт к образованию пировиноградной кислоты (пирувата), гликолиз в анаэробных условиях ведёт к образованию молочной кислоты (лактата).
Название «гликолиз» происходит от греч. γλυκός, glykos — сладкий и греч. λύσης, lysis — растворение.
Гликолиз — один из древнейших метаболических процессов, известный почти у всех живых организмов. Предположительно гликолиз появился более 3,5 млрд лет назад у первичных прокариотов.
Это процесс анаэробного распада глюкозы до образования ПВК, идущий с освобождением энергии. Гликолиз — общий начальный этап аэробного дыхания и всех видов брожения.
Локализация
Реакции протекают в цитоплазме и в хлоропластах и через промежуточные продукты могут взаимодействовать между собой. В цитозоле гликолитические ферменты находятся в составе мультиэнзимных комплексов, которые образуются с участием актиновых филаментов цитоскелета.
Общий обзор
Гликолитический путь представляет собой 10 последовательных реакций, каждая из которых катализируется отдельным ферментом.
Процесс гликолиза условно можно разделить на два этапа. Первый этап, протекающий с расходом энергии 2 молекул АТФ, заключается в расщеплении молекулы глюкозы на 2 молекулы глицеральдегид-3-фосфата. На втором этапе происходит НАД-зависимое окисление глицеральдегид-3-фосфата, сопровождающееся синтезом АТФ. Сам по себе гликолиз является полностью анаэробным процессом, то есть не требует для протекания реакций присутствия кислорода.
|
|
Результатом гликолиза является превращение одной молекулы глюкозы в две молекулы пировиноградной кислоты (ПВК) и образование двух восстановительных эквивалентов в виде кофермента НАД∙H.
Полное уравнение гликолиза имеет вид:
Глюкоза + 2НАД+ + 2АДФ + 2Фн = 2НАД∙Н + 2ПВК + 2АТФ + 2H2O + 2Н+.
При отсутствии или недостатке в клетке кислорода пировиноградная кислота подвергается восстановлению до молочной кислоты, тогда общее уравнение гликолиза будет таким:
Глюкоза + 2АДФ + 2Фн = 2лактат + 2АТФ + 2H2O.
Таким образом, анаэробном расщеплении одной молекулы глюкозы суммарный чистый выход АТФ составляет две молекулы, полученные в реакциях субстратного фосфорилирования АДФ.
Путь
Первой реакцией гликолиза является фосфорилирование молекулы глюкозы, происходящее при участии тканеспецефичного фермента гексокиназы с затратой энергии 1 молекулы АТФ; образуется активная форма глюкозы — глюкозо-6-фосфат (Г-6-Ф):
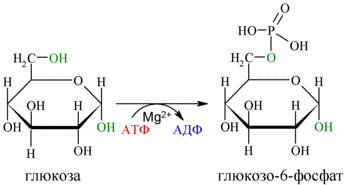
Для протекания реакции необходимо наличие в среде ионов Mg2+, с которым комплексно связывается молекула АТФ. Эта реакция необратима и является первой ключевой реакцией гликолиза.
Фосфорилирование глюкозы преследует две цели: во-первых, из-за того, что плазматическая мембрана, проницаемая для нейтральной молекулы глюкозы, не пропускает отрицательно заряженные молекулы Г-6-Ф, фосфорилированная глюкоза оказывается запертой внутри клетки. Во-вторых, при фосфорилировании глюкоза переводится в активную форму, способную участвовать в биохимических реакциях и включаться в метаболические циклы. Фосфорилирование глюкозы — это единственная реакция в организме, в которой глюкоза участвует как таковая.
|
|
В следующей реакции (2) ферментом фосфоглюкоизомеразой Г-6-Ф превращается во фруктозо-6-фосфат (Ф-6-Ф):

Энергия для этой реакции не требуется, и реакция является полностью обратимой. На данном этапе в процесс гликолиза может также включаться путём фосфорилирования и фруктоза.
Далее почти сразу друг за другом следуют две реакции: необратимое фосфорилирование фруктозо-6-фосфата (3) и обратимое альдольное расщепление образовавшегося фруктозо-1,6-бифосфата (Ф-1,6-бФ) на две триозы (4).
Фосфорилирование Ф-6-Ф осуществляется фосфофруктокиназой с затратой энергии ещё одной молекулы АТФ; это вторая ключевая реакция гликолиза, её регуляция определяет интенсивность гликолиза в целом.
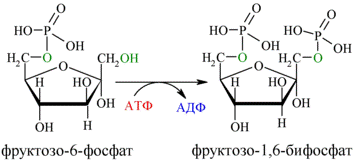
Альдольное расщепление Ф-1,6-бФ происходит под действием альдолазы фруктозо-1,6-бифосфата:
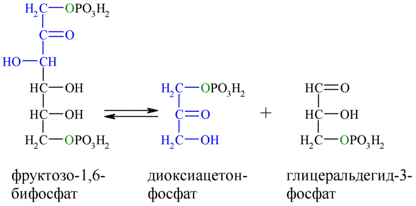
В результате четвёртой реакции образуются дигидроксиацетонфосфат и глицеральдегид-3-фосфат, причём первый почти сразу под действием фосфотриозоизомеразы переходит во второй (5), который и участвует в дальнейших превращениях:
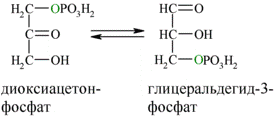
Каждая молекула глицеральдегидфосфата окисляется НАД+ в присутствии дегидрогеназы глицеральдегидфосфата до 1,3-дифосфоглицерата (6):
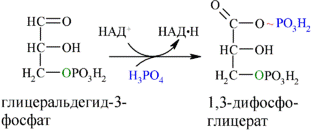
Далее с 1,3-дифосфоглицерата, содержащего макроэргическую связь в 1 положении, ферментом фосфоглицераткиназой на молекулу АДФ переносится остаток фосфорной кислоты (реакция 7) — образуется молекула АТФ:
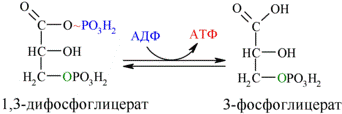
Это первая реакция субстратного фосфорилирования. С этого момента процесс расщепления глюкозы перестаёт быть убыточным в энергетическом плане, так как энергетические затраты первого этапа оказываются компенсированными: синтезируются 2 молекулы АТФ (по одной на каждый 1,3-дифосфоглицерат) вместо двух потраченных в реакциях 1 и 3. Для протекания данной реакции требуется присутствие в цитозоле АДФ, то есть при избытке в клетке АТФ (и недостатке АДФ) её скорость снижается. Поскольку АТФ, не подвергающийся метаболизму, в клетке не депонируется а просто разрушается, то эта реакция является важным регулятором гликолиза.
|
|
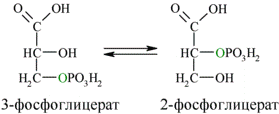
Затем последовательно: фосфоглицеролмутаза образует 2-фосфоглицерат (8):
Енолаза образует фосфоенолпируват (9):
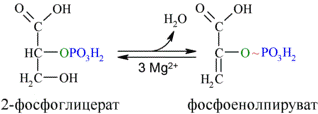
И наконец происходит вторая реакция субстратного фосфорилирования АДФ с образованием енольной формы пирувата и АТФ (10):
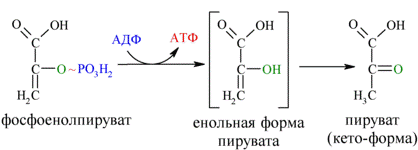
Реакция протекает под действием пируваткиназы. Это последняя ключевая реакция гликолиза. Изомеризация енольной формы пирувата в пируват происходит неферментативно.
С момента образования Ф-1,6-бФ с выделением энергии протекают только реакции 7 и 10, в которых и происходит к субстратное фосфорилировнаие АДФ.
Регуляция гликолиза
Различают местную и общую регуляцию.
Местная регуляция осуществляется путём изменения активности ферментов под действием различных метаболитов внутри клетки.
Регуляция гликолиза в целом, сразу для всего организма, происходит под действием гормонов, которые, влияя через молекулы вторичных посредников, изменяют
Регуляция гликолиза осуществляется через несколько ключевых этапов. Реакции, катализируемые гексокиназой (1), фосфофруктокиназой (3) и пируваткиназой (10) отличаются существенным уменьшением свободной энергии и являются практически необратимыми, что позволяет им быть эффективными точками регуляции гликолиза.
Регуляция гексокиназы
Гексокиназа ингибируется продуктом реакции — глюкозо-6-фосфатом, который аллостерически связывается с ферментом, изменяя его активность.
При фосфорилировании глюкоза теряет способность транспортироваться через мембрану молекулами-переносчиками, что создаёт условия для накопления её в клетке. Ингибирование гексокиназы Г-6-Ф ограничивает поступление глюкозы в клетку, предотвращая её чрезмерное накопление.
Глюкозо-6-фосфат может превращаться обратно в глюкозу при действии глюкозо-6-фосфатазы.
Регуляция фосфофруктокиназы
Интенсивность протекания фосфофруктокиназной реакции решающим образом сказывается на всей пропускной способности гликолиза, а стимуляция фосфофруктокиназы считается наиболее важным этапом регуляции.
Фосфофруктокиназа (ФФК) — это тетрамерный фермент, существующий поочерёдно в двух конформационных состояниях (R и T), которые находятся в равновесии и попеременно переходят из одного в другое. АТФ является одновременно и субстратом, и аллостерическим ингибитором ФФК.
Наиболее важным же аллостерическим регулятором гликолиза является фруктозо-2,6-бифосфат, который не является промежуточным звеном этих циклов. Фруктозо-2,6-бифосфат аллостерически активирует фосфофруктокиназу.
Синтез фруктозо-2,6-бифосфата катализируется особым бифункциональным ферментом — фосфофруктокиназой-2/фруктозо-2,6-бифосфатазой (ФФК-2/Ф-2,6-БФаза). В нефосфорилированной форме белок известен как фосфофруктокиназа-2 и имеет каталитическую активность по отношению к фруктозо-6-фосфату, синтезируя фруктозо-2-6-бифосфат. В результате чего значительно стимулируется активность ФФК и сильно ингибируется активность фруктозо-1,6-бифосфатазы. То есть при условии активности ФФК-2 равновесие этой реакции между гликолизом и глюконеогенезом смещается в сторону первого — синтезируется фруктозо-1,6-бифосфат.
В фосфорилированном виде бифункциональный фермент не обладает киназной активностью, а наоборот в его молекуле активируется сайт, который гидролизует Ф2,6БФ на Ф6Ф и неорганический фосфат. Метаболический эффект фосфорилирования бифункционального фермента состоит в том, что аллостерическая стимуляция ФФК прекращается, аллостерическое ингибирование Ф-1,6-БФазы ликвидируется и равновесие смещается в сторону глюконеогенеза. Продуцируется Ф6Ф и затем — глюкоза.
Пируваткиназа
Следующим шагом, где осуществляется регуляция гликолиза, является последняя реакция — этап действия пируваткиназы. Для пируваткиназы также описан ряд изоферментов, имеющих особенности регуляции.
Блокируют процесс гликолиза ингибиторы SH-содержа-щих ферментов (например, дегидрогеназы ФГА) — иод- или фторсодержащие соединения.
Характеристика гликолиза:
- большинство реакций обратимо, за исключением трех (реакций 1, 3, 10);
- все метаболиты находятся в фосфорилированной форме;
- источником фосфатной группы в реакциях фосфорилирования являются АТР (реакции 1, 3) или неорганический фосфат (реакция 6);
- регенерация NAD+, являющаяся необходимым условием протекания гликолиза, происходит при аэробном гликолизе посредством дыхательной цепи. В этом случае водород транспортируется в митохондрии с помощью челночного механизма при участии переносчиков. Это происходит потому, что мембрана митоходрий непроницаема для протонов. При анаэробном гликолизе регенерации NAD+ осуществляется независимо от дыхательной цепи. В этом случае акцептором водорода от NADH является пируват, который восстанавливается в лактат;
- образование АТР при гликолизе может идти двумя путями: либо субстратным фосфорилированием, когда для фосфорилирования ADP используется энергия макроэргической связи субстрата (реакции 7, 9), либо путем окислительного фосфорилирования ADP, сопряженного с дыхательной цепью (реакция 6).
В растительных клетках помимо «прямого» пируватки-назного пути осуществляется «обходной» путь превращения ФЕП с помощью ФЕП-карбоксилазы. Этот цитозольный энзим играет важную роль в Сз-растениях и в нефотосинтезирующих тканях С4- и САМ-растений, направляя интермедиаты дыхания в реакции биосинтеза.
Возможно обращение гликолиза благодаря обратимости действия большинства его ферментов. Необратимы лишь реакции фосфорилирования глюкозы и фруктозы, а также реакция образования пирувата из ФЕП. Первые две реакции, катализируемые гексо- и фруктокиназами, могут быть обращены с помощью других ферментов — фосфатаз, которые осуществляют дефосфорилирование. Обходной путь превращения пирувата в фосфоенолпируват сложнее. Он идет через оксалоацетат, который образуется из пирувата в митохондриях. После прохождения через мембрану митохондрий пируват карбоксилируется пируваткарбоксилазой (ПДК) до оксалоацетата, который восстанавливается НАД-зависимой митохондриальной МДГ до мала-та. Последний переносится в цитоплазму, где вновь превращается в оксалоацетат цитоплазматической НАД-зависимой МДГ. В цитоплазме возможно и прямое превращение пирувата в малат (восстановительное карбоксилирование), но вклад этого процесса невелик. Далее оксалоацетат под действием ФЕП-карбоксикиназы и с участием АТФ декарбоксилируется и образуется ФЕП. Реакции обращения гликолиза включены в процесс образования глюкозы из неуглеводиых предшественников, т. е. глюконеогенез. Обращение гликолиза и глюконео-генез упрощаются при участии ФФн-ФФК в обратимой реакции образования фруктозобисфосфата.
Значение
Гликолиз — катаболический путь исключительной важности. Он обеспечивает энергией клеточные реакции, в том числе и синтез белка. Промежуточные продукты гликолиза используются при синтезе жиров. Пируват также может быть использован для синтеза аланина, аспартата и других соединений. Благодаря гликолизу производительность митохондрий и доступность кислорода не ограничивают мощность мышц при кратковременных предельных нагрузках.
Таким образом, гликолиз выполняет две фундаментальные функции: окисление гексоз для генерации АТФ, восстановителей, пирувата и синтез гексоз из низкомолекулярных соединений в процессе энергозависимого глюконеогенеза.
Энергетический выход. При окислении одной молекулы глюкозы образуются две молекулы ПВК. При этом за счет первого и второго субстратного фосфорилирования, т. е. фос-форилирования, происходящего непосредственно в субстрате в результате внутримолекулярных перестроек, образуются четыре молекулы АТФ. Однако две молекулы АТФ тратятся на фосфорилирование глюкозы и фруктозо-б-фосфата. Поэтому чистый выход гликолитического субстратного фосфорилирования составляют две молекулы АТФ. При использовании в качестве субстрата сахарозы и при участии в гликолизе ФФн-ФФК энергетический выход увеличивается вдвое, так как АТФ-ФФК заменяется ФФд-ФФК и затрат АТФ на образование глюкозо-1-фосфата в инвертазной реакции и на активирование фруктозо-6-фосфата не происходит.
Помимо того, в реакции окисления ФГА до ФГК на каждую из фосфотриоз восстанавливается по одной молекуле НАДН. Окисление же каждой молекулы НАДН в ЭТЦ митохондрий сопряжено с синтезом трех молекул АТФ. В расчете на две триозы, т. е. на исходную молекулу гексозы это составляет шесть молекул АТФ. Следовательно, в целом энергетический выход гликолиза равен восьми (или 10) молекулам АТФ. Поскольку свободная энергия гидролиза одной молекулы АТФ в клетке составляет 41,868 кДж/моль (10 ккал), восемь (или 10) молекул АТФ дают 335 (или 419) кДж/моль (80-100 ккал).