Известно, что клетки прокариот размножаются бесполым путем, простым бинарным делением. У многоклеточных эукариот некоторые соматические клетки также могут делиться простым делением, перетяжкой ядра и цитоплазмы. В чем принципиальная разница этих двух типов размножения клеток? А чем от этих типов деления отличается бесполое размножение одноклеточных эукариот?
Решение. У многоклеточных эукариот некоторые соматические клетки могут делиться простым делением, при которой возникает перетяжка ядра и цитоплазмы. Оно происходит без образования веретена деления и равномерного распределения хромосом, а также без характерной для митоза спирализации хромосом и называется амитозом. Неизвестно синтезируется ли перед началом амитоза ДНК и как происходит распределение ДНК между дочерними ядрами. Одно из возможных объяснений: по крайней мере в ряде случаев сегрегация генома осуществляется путём перешнуровывания полиплоидного ядра, не приводящее к обязательному строго равномерному расхождению сестринских хроматид, как при митозе. После амитоза клетка не способна вернуться в нормальный митотический цикл. В норме у человека наблюдается в высокоспециализированных тканях, в клетках, которым уже не предстоит делиться митотически – в эпителии и печени.
Деление же бактериальных клеток потенциально может происходить неограниченное количество раз. При этом в бактериальных клетках происходит строго закономерная сегрегация идентичных молекул ДНК бактериальных хромосом (но не всегда плазмид), которая не отделена от цитоплазмы мембраной (то есть не находится в ядре, как у эукариот). Одна из традиционных моделей сегрегации бактериальных хромосом основана на представлении о заякоривании ДНК на мембране и участии мембраны в сегрегации. Однако в настоящее время предложен уже целый ряд разнообразных моделей сегрегации бактериальных хромосом при делении, каждая из которых применима к определённым систематическим группам (см. Пиневич «Микробиология. Биология прокариотов. Том 3»
По современным представлениям при делении бактериальных клеток, в отличие от деления эукариотических клеток, ДНК может быть недореплицирована и, таким образом, в дочерних клетках иметь дробную копийность (рисунок 1).
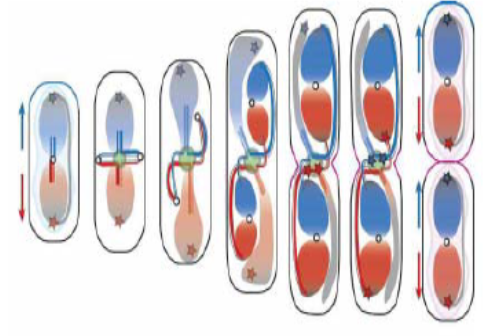
Рисунок 1. Сегрегация бактериальных хромосом при делении клеток в недореплицированном состоянии. Красным и синим цветами показаны идентичные копии реплицированных молекул ДНК.
Бесполое же размножение эукариот обязательно происходит с помощью митоза с участием митотического веретена, нити которого прикрепляются к центромерам и осуществляют сегрегацию сестринских хроматид. Митоз при этом может происходить без растворения ядерной оболочки, как, например, у эвгленозой и грибов (закрытый митоз). В этом случае веретено деления образуется внутри ядра. При открытом митозе ядерная оболочка исчезает и хромосомы с веретеном деления оказываются в метафазе и анафазе митоза в цитоплазме.
Биологический смысл полового размножения – обмен генетическим материалом и возникновение диплоидности, что дает возможность корректировки возникающих ошибок в генетическом «тексте». Как же прокариотам, у которых половое размножение отсутствует, а геном гаплоиден, удается в течение миллиардов лет не вымереть от неизбежно накапливающихся в геноме ошибок?
Решение. Копирование генетической информации обязательно происходит с теми или иными ошибками. Если у организмов без рекомбинации разных геномов возникает вредная мутация, то потомки уже не смогут от неё избавиться. Поэтому число вредных мутаций в таком организме может быть либо постоянным путём более сильного отбора, либо увеличиваться (храповик вращается только в одну сторону). Если в сообществе бесполых особей случайно погибает особь с наименьшим количеством вредных мутаций, то потеря необратима (храповик Мёллера). Таким образом, если бы организмы размножались только самовоспроизведением без рекомбинации, то они быстро бы накопили груз вредных мутаций.
У бактерий существует большое разнообразие механизмов рекомбинации, позволяющих избавиться от действия храповика Мёллера.
В основе этих механизмов лежит один фундаментальный факт: если в бактериальную клетку проникнет фрагмент ДНК, гомологичный участку хромосомы, то с высокой частотой происходит гомологичная рекомбинация.
Именно поэтому в бактериальной хромосоме практически не встречаются длинные (более 100 п.н.) повторяющиеся фрагменты ДНК.
1. Конъюгация - перенос генетического материала от одной бактериальной клетки (донора) к другой (реципиенту) при их непосредственном контакте. Контакт клеток может осуществляться т.н. пилями (у грамотрицательных) или непосредственно. Генетический материал может переноситься с помощью специальной плазмиды (например, у E.coli так называемая F-плазмида может интегрировать в хромосому, как изображено на рисунке 2). При её переносе реципиентную клетку также будет происходить перенос хромосомной ДНК. Рекомбинантная клетка станет мерозиготой (неполная диплоидность).

Рисунок 2. Схема переноса части бактериальной хромосомы с помощью F-плазмиды при конъюгации у E.coli.
Наличие повторяющихся фрагментов хромосом включает механизм гомологичной рекомбинации. F-фактор ведет себя как геномный паразит, а конъюгация и f-пили несомненно являются всего лишь средствами передачи инфекции.
На этой основе и возник полезный для самой бактерии половой процесс; возможно, способность F-фактора встраиваться в бактериальную хромосому была поддержана естественным отбором.
Перенос части хромосом с помощью конъюгативных плазмид встречается лишь у небольшой части бактерий.
Встречается у десятков родов бактерий: энтеробактерий, псевдомонад и др.
Возможна конъюгация между разными «видами» бактерий, например, кишечной палочкой и сальмонеллой, что объясняется родственными связями между этими таксонами и большой долей гомологичной ДНК.
Наряду с конъюгативными плазмидами в рекомбинации могут участвовать конъюгативные транспозоны. В отличие от плазмид конъюгативные транспозоны не содержат точки ori.
Обнаружены у представителей родов Bacillus, Clostridium, Escherichia, Neisseria, Salmonella, Pseudomonas и др. (более 50 родов).
Важно, что нередко с конъюгативными плазмидами и транспозонами одновременно переносятся большие фрагменты хромосом.
Однако возможно, что у большинства видов конъюгация все же отсутствует.
2. Трансдукция – перенос генетического материала у прокариот с помощью фагов. При сборке в фаговые частицы иногда попадает не фаговая ДНК, а ДНК бактерий. Попадая в другую бактериальную клетку, фрагмент ДНК может включаться в её геном, обычно путём гомологичной рекомбинации.
3. Горизонтальный перенос генов (ГПГ), или трансформация – непосредственный захват генетического материала бактериальной клеткой из окружающей среды. Одна из основ горизонтального захвата генов между даже удалёнными формами, что резко ускоряет эволюцию.
Гомологичная рекомбинация становится возможной благодаря тому, что бактерии захватывают родственный генетический материал. Наряду с этим ГПГ обеспечивает захват чужеродного генетического материала, что важно как способ быстро эволюционировать и лучше приспособиться к окружающим условиям (рисунок 3).
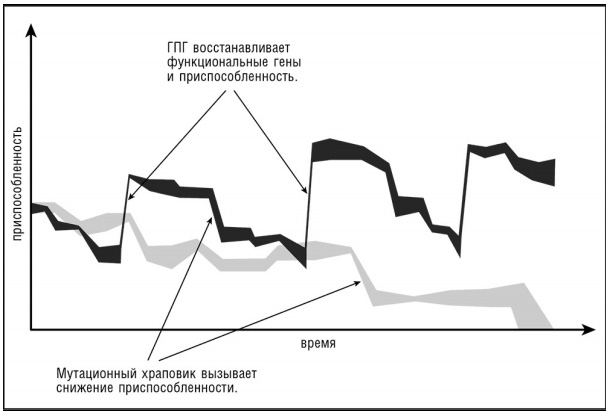
Рисунок 3. Роль ГПГ в избавлении от храповика Мёллера и повышении приспособленности.
Многие бактериальные клетки имеют приспособления для преимущественного захвата лишь той ДНК из окружающей среды, которая образована близкородственными штаммами.
Возможно самый распространённый механизм такого ограничения – системы рестрикции-модификации.
Рестриктаза расщепляет чужеродную ДНК по определённому сайту узнавания (например, EcoRI расщепляет двуцепочечный сайт 5’-GAATTC-3’). Собственная ДНК клетки не расщепляется рестриктазой, так как метилаза метилирует основание в сайте узнавания (рисунок 4).

Рисунок 4. Принцип функционирования системы рестрикции-модификации: рестриктаза расщепляет чужеродную неметилированную последовательность (а), но не может расщепить собственную ДНК, «меченую» метильной группой СН3- по сайту узнавания соответствующей метилазы (b).
Таким образом, система рестрикции модификации помогает осуществлять преимущественно рекомбинацию только между близкородственными штаммами.
Существуют и другие менее распространённые способы распознавания преимущественно своей ДНК, например, наличие коротких повторов, узнаваемых мембраной, как это происходит, например, у Neisseria gonorrhoeae.
Бактерии выработали специальные механизмы попадания бактериальной ДНК в раствор:
1) Лизис части клеток, включаемый при определённых условиях, например, при достижении бактериальной популяции определённой плотности (например, у Bacillus subtilis).
2) Фаги с определённой частотой (не излишне высокой благодаря системе рестрикции модификации) осуществляют периодический лизис клеток. Таким образом, фаги способствуют рекомбинации бактериальной ДНК как через трансдукцию, так и через трансформацию (рисунок. 5).
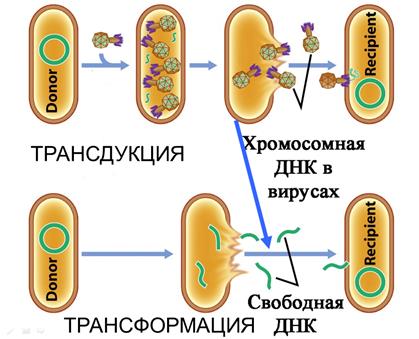
Рисунок. 5. Механизмы рекомбинации ДНК, обеспечиваемые фагами: трансдукция (прямой перенос в вирусных частицах) и трансоформация (опосредованный перенос через ДНК, оказавшуюся вне клетки благодаря лизису, индуцируемому фагом).
Репарация повреждённой ДНК также отчасти возможна благодаря тому, что у большинства бактерий хромосома представлена не в единственном экземпляре (например, у E.coli обычно 4-8, у цианобактерий – 8-12 и т.д.). В частности, Deinococcus radiodurans способен выдерживать высокие дозы радиации благодаря тому, что в его клетках присутствует сразу четыре копии генома. "Запасные" копии нужны этому обитателю ядерных реакторов для оперативной репарации (починки) ДНК. Когда под воздействием радиации в одной из хромосом возникают мутация или разрыв, запасные хромосомы, по-видимому, используются как матрицы, с которых можно скопировать "правильные" последовательности нуклеотидов в испорченную хромосому.
Что называют чередованием поколений? Есть ли чередование поколений цветковых растений? А у млекопитающих? Ответ аргументируйте.
Чередование поколений - периодическая смена двух или более поколений особей одного вида, различающихся между собой генетически (например, плоидностью), анатомо-морфологически, физиологически, поведенчески и способом размножения. При чередовании поколений чаще одно или несколько бесполых поколений организмов сменяется поколением организмов, размножающихся половым путем.
У цветковых растений есть чередование поколений. Половое поколение представлено гаплоидным гаметофитом, бесполое — диплоидным спорофитом. Споры у цветковых растений прорастают прямо внутри спорофита. Внутри семязачатка макроспора прорастает в женский гаметофит, в нём образуется яйцеклетка. Внутри пыльцевого мешка микроспора превращается в мужской гаметофит – пыльцевое зерно, в нем образуются спермии. Женский гаметофит цветковых растений называется зародышевый мешок. Он содержит 7 клеток, в том числе яйцеклетку и центральную диплоидную клетку.
Мужской гаметофит цветковых состоит из двух клеток – вегетативной (превращается в пыльцевую трубку) и генеративной (превращается в два спермия).
Оплодотворение у цветковых двойное:
1) один спермий сливается с яйцеклеткой, получается диплоидная зигота, которая превращается в зародыш спорофита,
2) второй спермий сливается с диплоидной клеткой, получается триплоидный эндосперм.
У млекопитающих чередования поколений нет, поскольку гаплодиные клетки представлены только гаметами (рисунок 6). Многоклеточный организм развивается на диплоидной фазе.

Рисунок 6. Схема жизненного цикла млекопитающих. Рисунок взят из статьи Д. Шабанова «Чудеса полового размножения». «Компьютерра», https://old.computerra.ru/own/shabanov/663786/
Могут ли существовать организмы, размножающиеся половым путем, но у которых отсутствует мейоз? Какими особенностями должны обладать такие организмы? Существуют ли они в природе?
Решение. Мейоз в классическом понимании – это последовательность двух делений ядра диплоидных клеток, где в результате первого деления после рекомбинации в профазе расходятся гомологичные хромосомы, а в результате второго деления – сестринские хроматиды.
Настоящий мейоз может отсутствовать, если есть другие механизмы уменьшения в два раза числа плоидности клеток. Такие механизмы найдены у грибов (например, пеницилла и аспергилла) при так называемом парасексуальном процессе. Гифы разных генетических индивидов соприкасаются и передают друг другу ядра. Разные ядра могут сливаться, образуя гибридные ядра с двойным набором хромосом. Эти ядра, как и остальные, делятся надвое митотически. Однако при митозах гибридных ядер у грибов в них шаг за шагом происходит элиминация (удаление) избыточных хромосом. Иногда при этом наблюдается рекомбинация — обмен участками между парными хромосомами, образование новых сочетаний генетического материала. Разные ядра, образовавшиеся после упрощения гибридного ядра, имеют разные по происхождению части хромосом. Возможно в парасексуальном процессе важную роль играет митотическая рекомбинация. На рисунке 7 гифы, где ядра имеют гаплоидный (единичный) набор хромосом, показаны серым, а диплоидные (с двумя хромосомными наборами) окрашены зеленоватым.

Рисунок 7. Упрощённая схема парасексуального процесса у грибов. Рисунок взят из статьи Д. Шабанова «Чудеса полового размножения». «Компьютерра», https://old.computerra.ru/own/shabanov/663786/
Таким образом, диплоидные ядра грибов становятся гаплоидными постепенно, после многих митозов, а в одних и тех же гифах могут находиться различные ядра.